
- 1Spark基础05-map和mapValue_spark mapvalues
- 2一分钟看懂IDEA中的GIT LOG视图
- 3AES算法的CBC和ECB两种工作模式_ecb加密密钥要求
- 4第十三届蓝桥杯省赛真题 Java B 组【原卷】_蓝桥杯13届javab组
- 5使用ollama本地跑大模型_ollama问题
- 6解决 Android Studio 创建项目时极其的慢的尴尬_android studio 建项目慢
- 7探索未来飞行:HKUST-Aerial-Robotics 的 Fast-Planner 项目
- 8AI实战 | AI算法能不能协助英语老师出一份nb的考卷?
- 9相似向量检索库-Faiss-简介及原理
- 10MySQL与NoSQL之间的区别
意识理论综述:众多竞争的意识理论如何相互关联?
赞
踩
来源: 集智俱乐部
作者:Anil K. Seth & Tim Bayne
译者:李路凯、陈斯信
编辑:邓一雪
导语
意识和底层的神经活动之间如何联系起来?试图解释这一问题的意识理论层出不穷。然而,随着实验数据积累,众多的意识理论并没有被逐渐证伪,反而数量持续增加,这种现象促使人们对现有理论进行整合。2022年5月发表于 Nature Reviews Neuroscience 的综述文章,回顾了四类重要的「意识理论」:高阶理论、全局工作空间理论、整合信息论和复馈/预测处理理论。通过经典的实验争论说明这些理论的联系与区别,从而为理解意识科学的研究现状提供了一个视角,以期推动意识理论和实验研究之间的良性循环,使我们更深刻地理解意识这个最深刻的奥妙。
研究领域:意识理论,认知神经科学

论文题目:
Theories of consciousness
论文链接:
https://www.nature.com/articles/s41583-022-00587-4
目录
摘要
一、引言
二、预备知识
三、四类意识理论路径
1. 高阶理论
2. 全局工作空间理论
3. 整合信息论
4. 复馈/预测处理理论
四、意识理论的评估标准和发展方向
五、展望
摘要
近年来,有关意识的生物和物质基础的理论层出不穷。好的理论能够指导实证研究,解释实验数据,开发新型实验技术,以及扩展我们控制感兴趣的实验现象的能力。然而,在意识领域,目前的理论如何相互关联或者能否用实验鉴别,尚不清楚。为了阐明这一复杂的领域,我们回顾了四类重要的意识理论(Theories of Consciousness,TOC)路径:高阶理论(higher-order theories,HOT)、全局工作空间理论(global workspace theories,GWT)、整合信息论(integrated information theory,IIT)和复馈/预测处理理论(re-entry and predictive processing theories)。通过指明它们试图解释的意识问题、它们的神经生物学承诺、以及它们引证的实验证据,刻画了这四类路径的主要特征。我们考虑了一些重要的实验争论如何可能区分不同理论,并列举了三个意识理论需要发展的方向,使它们能够被神经科学实验所检验。有理由相信,意识理论的迭代发展、验证和比较将导向更深刻的理解。
一、引言
在意识科学复苏的最初几十年,研究者们侧重于寻找“意识的神经相关物” (neural correlates of consciousness, NCC)。严格地说,NCC是足以构成某种意识状态的最小神经事件集;在实践中,寻找NCC包括寻找与意识最密切相关的大脑状态和过程[1-3]。专注于寻找NCC的工作是有用的,因为 NCC“理论中立”,为具有不同理论和哲学背景的研究人员提供了共同的语言和方法。然而,NCC框架的局限性变得越来越明显,比如难以将NCC从意识的神经先决条件和附带影响中区分出来[4-7]。为了应对这些限制,人们越来越关注意识理论(theories of consciousness, ToC)的发展。在ToC的帮助下,我们能超越基于NCC的方法,转而去探索具有解释性洞见的意识模型。事实上,意识科学的主要目标就是得到被实验验证的ToC[8,9]。
NCC的研究方法强调意识与大脑活动之间的相关性,而ToC则侧重于鉴别意识特征与神经机理之间的解释性联系[10]。话虽如此,理论家们经常采用不同的话语体系来解释神经活动和意识之间的联系。一部分人认为,令人满意的ToC应该缩小“解释鸿沟”(方框 1),使神经活动和意识之间的关系,像水的化学结构与其总体特性之间的关系一样,清晰明了[11]。另一部分人认为,“解释鸿沟”不一定能完全消除,但仍然希望有一个框架可以解释意识的某些方面,减少或消除有关意识的生理基础的神秘感[12,13]。还有一部分人认为, “解释鸿沟”具有误导性,不应该被意识科学认真对待[14,15]。
方框1
意识理论和“困难问题”
20世纪90年代,大卫·查默斯 (David Chalmers) 区分了意识的“困难”和“简单”问题[164]。简单问题涉及与意识相关的功能和行为,而困难问题涉及意识的(现象,主观)体验。使这个难题变得困难的是“解释鸿沟”[165]——这是一种直觉,即似乎不可能用物理或功能术语对主观体验给予完全还原论的解释。
一些意识理论(比如整合信息论和某些版本的高阶理论)直接着手处理困难问题。其它一些理论(比如全局工作空间理论)关注与意识相关的功能和行为特性,尽管可以认为它们也着手处理了困难问题,但这并不是其支持者的主要目标。第三种策略(被一些预测处理理论家采用)旨在提供一个框架,在其中可以解决有关意识现象特征的各种问题,而不试图解释现象本身的存在性[67]——这有时被称为“真正的问题”[13, 166]。
意识领域的一个关键问题是,困难问题应当由科学来解决吗?还是应当被搁置而不是被解决?认为困难问题应当被搁置的人经常争辩说,正是我们用来表达自身意识状态的概念(“现象概念”)的特殊性,导致了这个难题的出现[167,168]。一个相关的观点是幻觉主义,它认为我们实际上并没有现象状态,而只是将自己表示为具有这种状态[14,15]。不管这些假说的优点是什么,随着我们解释、预测和控制意识的现象属性和功能属性的能力增强,困难问题或许会变得没那么棘手[166,169]。
表1列出了现有的众多ToC。值得注意的是,随着实验数据的积累,众多ToC并没有逐渐被证伪从而被放弃,它们的数量不降反增。这种现象促使人们将现有理论相互整合[16],或进行“对抗式合作”。在“对抗式合作”中,对于某个实验的结果是否会支持或破坏某个理论,不同理论的支持者们需要事先达成一致[17]。然而,理论整合和“对抗式合作”都存在重大挑战,我们将讨论这一点。
表格1 | 意识理论选集
理论 | 核心观点 | 关键文献 |
高阶理论 | 对低阶心智状态的高阶表征产生意识 | 31,46 |
自组织高阶表征理论 | 意识是大脑对自己的高阶表征 | 34,140 |
受注意的中间表征理论 | 被注意力放大的中间层表征产生意识 | 141,142 |
全局工作空间理论 | 点火并广播到神经元全局工作空间的信息进入意识,额叶与顶叶承担中心枢纽般的作用 | 47–49 |
整合信息理论 | 意识就是产生最大不可约整合信息的物质基础的因果结构 | 57,59,60 |
闭合信息理论 | 意识依赖于对环境的非平凡信息闭合,特别是粗粒度水平 | 143 |
动态核心理论 | 神经活动的功能集群结合了高层次的动态整合与分化,产生意识 | 144 |
神经元达尔文主义 | 自然选择原理塑造的价值依赖型学习的历史信息,反应在复馈行为上,产生意识 | 145,146 |
局部循环 | 意识依赖于皮层的局部复馈或循环过程,能促进学习 | 65,71 |
预测处理 | 感知依赖于对感觉信号的成因的预测推断。它提供了一个框架,有助于系统地将神经机制映射到意识的各个方面 | 67,73,79 |
神经元表征理论 | 意识依赖于多级神经编码的预测表示 | 84 |
主动推断 | 具有不同的观点,比如意识依赖于对自主行为的时序的和反事实的深度推理 | 76;另见91 |
野兽机器理论 | 以意识依赖于以内稳态控制为导向的预测推理 | 13,75,77;另见90 |
神经元主观架构 | 意识依赖于提供第一人称视角身体状态的神经图谱 | 24 |
自我浮现理论 | 意识依赖于内稳态程序和多层次内感受图之间的相互作用,以情感和感觉为核心 | 23,147 |
注意力图式理论 | 意识依赖于注意力控制的神经编码模型 | 148 |
多重草稿模型 | 意识依赖于多个(可能不一致的)表示,而不是中央系统可用的单一、统一的表示 | 149 |
感觉运动理论 | 意识依赖于对感觉运动突发事件规律的掌握 | 88 |
无限联想学习 | 意识依赖于一种学习形式,使生物体将自身的驱动力与新颖、复合和非反射诱导的刺激或动作联系起来 | 150 |
树突整合理论 | 意识依赖于细胞水平的自上而下和自下而上信号的整合 | 151 |
电磁场理论 | 意识就是编码在大脑电磁场中的物理整合的、因果活跃的信息 | 152 |
调谐客观还原理论 | 意识依赖于神经元内的微管中发生的量子计算 | 18 |
在这篇综述中,我们考虑了ToC之间的关系,以及它们与实验数据的联系。我们指明了几条发展路径,能够让理论发展和实证研究相互支持和促进,从而寻找令人满意的对意识体验的科学解释。我们只考虑那些本身以神经生物学术语表达的理论,或者被合理地认为包含可以用神经生物学术语表达的主张的理论。(正如我们将看到的,一些“神经生物学”ToC使用信息论或“功能之间的联系”等抽象语言进行表达,仅因为其抽象语言能与特定的神经机制建立联系而被称为“神经生物学”)。另外,我们只考虑与已知物理理论一致的神经科学理论。我们也不考虑将意识直接与量子力学过程联系起来的理论(比如[18,19])。
二、预备知识
ToC之间“鸡同鸭讲”的主要原因之一是它们通常具有不同的解释目标。因此,我们首先考虑,一个详尽的ToC需要解释哪些内容。即使这个问题也存在争议,理论家们在ToC应该解释什么样的意识现象上经常无法达成一致。
意识的核心问题是“第一人称体验”或“主观知觉”。虽然,除了循环定义以外,无法对这些术语进行定义,但意识研究的对象可以用一些直观的特征来进行区分和限定。比如使用“有某种感觉”(something it is like)来描述某个生物体具有意识[20],使用“感觉起来怎么样”(what it is like)来区分两种不同的意识状态。一个详尽的ToC不仅需要解释为什么一些生物体或系统有意识而另一些没有意识,还需要解释为什么不同的过程导致不同的意识状态。
意识状态可以分为两类:全局状态和局部状态。全局状态描述生物体的整体主观特征,并与觉醒水平的变化、行为反应的变化有关。常见的全局状态包括清醒、做梦、镇静、最低意识状态(或许还包括迷幻状态)。这些全局状态有时被称为“意识水平”,但我们更喜欢“全局状态”这个术语,因为它留下了这样一种可能性,即这些状态无法在单一维度上被完整地排序,而最适合在多维空间内被描述[21]。
局部状态——通常被称为“意识的内容”或具有“感质(qualia)”的状态——的特征被“这种状态感觉起来怎么样”所刻画。头痛的局部状态与闻咖啡的局部状态不同,因为头痛的感觉与闻咖啡的感觉不同。局部状态的描述可以具有多层级的颗粒度,从低级的感知特征(例如颜色)到单个物体,再到完整的多模态场景感知。一些重要的局部状态构成了自我体验的基础,自我体验包括情绪、情感、意志、拥有身体的感觉、明晰的自传体记忆等[13, 22-24]。虽然神经生物学理论倾向于关注具有感觉和感知内容的局部状态,但意识还包括具有认知和命题内容的局部状态,例如解决填字游戏时出现的想法。重要的是,在特定的时刻,一个生物体具有的众多局部状态并非简单而独立地出现,相反,它们作为单一的意识场景的组成部分,被绑定在一起[25, 26]。
第二个区别是意识的现象属性和功能属性之间的区别。现象属性是指意识的主观体验特征,也就是说“感觉起来怎么样”。功能属性关注的是,对具有意识并因此获得好处的生物体而言,在它的认知经济性方面,心智状态承担了什么角色。(这里的“功能”包括目的论功能——由进化形成的功能——和部署性功能——一个过程在它所属的更大系统的运作过程中所起的作用;见参考文献 27)例如,有意识地看到咖啡杯可以实现一系列功能,包括灵活地使用杯子(可能是从杯子里喝水,或者把它扔到房间的另一头),对上述事件进行情景记忆,以及提供有关上述体验的口头报告。虽然我们区分了意识的现象属性和功能属性,但并不是说它们相互独立(它们很可能不是独立的),只因为它们为 ToC 提供了不同的解释目标。正如我们将看到的,一些 ToC 关注意识的现象特征,另一些则关注意识的功能特征,还有一些试图同时解释意识的功能特征和现象特征。
第三个区别是ToC可能试图回答的,有关局部状态(“内容”)的两种问题。一方面,人们可能会问为什么生物体处于某个局部状态(而不是另一个局部状态)。另一方面,人们可能会问,为什么一个特定的局部状态具有它所具有的主观体验特征(而不是其它类型的特征)。这种区别可以用双目竞争实验来解释。在双目竞争实验中,被试的每只眼睛都接收到不同的刺激(例如,右眼呈现房子,左眼呈现人脸),被试的视觉体验在左眼刺激和右眼刺激之间交替[28]。在特定时刻,意识内容只包含房子,而不包含人脸,此时,我们可以问,为什么在这一刻对应于“房子”的心理状态是有意识的(而“人脸”的心理状态无意识),我们也可以问为什么房子的视觉体验具有它们所具有的独特的体验特征,而不是具有看到一张脸,听到铃铛响或感到疼痛时的体验特征。值得注意的是,可能有一些内容是不可能有意识的(例如,初级感知或调节系统中的低级处理过程),而其它内容只能是有意识的(例如,全局整合的感知场景)。因此,除了解释为什么某些心理内容在某些情景中是有意识的,而在其它情景中不是,另一个挑战是解释,为什么某些内容永远不会是有意识的,而某些内容只能作为有意识的存在。
大多数ToC并没有试图同时解决我们刚刚阐明的所有意识问题,它们试图解释意识的某些方面的问题,它们或许将这种努力看做迈向详尽完整的意识理论的一步。虽然,只研究意识的某一方面,这种局限本身并不构成对某一ToC的批驳和反对,但这确实意味着意识理论之间的相互比较变得没有那么简单而直接了。如果不同理论针对意识的不同方面(例如,一种理论专注于意识的现象特征,另一种理论关注其功能特征),那它们乍一看可能不像“对手”。
本文回顾的ToC分为四类:高阶理论(HOT),全局工作空间理论(GWT),整合信息论(IIT)以及复馈/预测处理理论。虽然某些对意识的解释跨越了多个类别,而另一些则无法合理地归入这些类别中的任何一个(表格 1),但这种四分类方法提供了一个视角,有助于理解意识科学的研究现状(方框2;关于其它对理论进行分组的方式,见[29])。在下文中,我们将介绍每个类别的关键元素,描述一些值得注意的类别内差异,并指明最密切相关的意识问题。然后,我们根据一些经典的实验争论,来说明这些ToC如何相互关联,并提出了几个建议,我们认为这些建议将有助于推动意识的理论发展和实验研究之间的良性循环。
方框2
其他方案:注意、学习和情感
除了本文提及的理论方法(表1)之外,意识理论还包含许多其它方向。一种方法侧重于注意力。例如,Graziano的注意力图式理论(attention schema theory)将有意识的感知与注意力控制模型联系起来[148]。另一种基于注意力的意识理论是被注意的中间表征理论(attended intermediate representational theory)。该理论首先由Jackendoff[141]提出并由Prinz[142]发展,该理论认为,当中级感知表征获得注意力时,意识就会发生。
其它理论方向侧重于学习。比如Jablonka和Ginsburg认为,“无限联想学习”提供最小意识,这种学习机制使生物体将自身的驱动力与新颖、复合和非反射诱导的刺激或动作联系起来[150]。其它基于学习的理论与我们已经描述的一些理论部分重叠,比如Cleeremans版本的高阶理论[34,140]和Lamme的局部循环理论(local recurrency account),后者认为循环信号凭借其在学习中的作用产生意识[65]。基于学习的理论也与“选择主义”密切相关,它们认为意识来源于神经元团簇内部和之间的类似进化的动力学[145,146]。
基于情感的理论强调大脑在生理调节中的作用,作为意识的基础。比如Damasio认为意识依赖于生物体生理状况的分层嵌套表征[147,170],他还将情感与预测处理过程相结合,在控制导向的内感受预测过程中建立意识体验[13,77,90]。一些基于情感的理论否认皮层中的机制是意识所必需的,而将产生意识的机制定位在脑干中[171,172](反对的观点另见参考文献 173)。
三、四类意识理论路径
1. 高阶理论
所有高阶理论(HOT)共有的核心主张是,某种精神状态是有意识的,当且仅当它成为某种元表征状态指向的目标。元表征不仅仅指在层次化处理结构中位于更高层或更深层的表征,而是指以其它表征为指向目标的表征(图1)。例如,内容为“我正在经历的视觉体验中包含一个正在移动的点”的表征是一个元表征,因为它的内容涉及自己对世界的表征,而不是世界本身。
不同HOT之间的重要区别之一,是它们对负责意识的元表征的性质和作用的解释不同。某些HOT发现某些元表征对包含思想的意识(或类似思想的状态)至关重要[30-32]。另一些HOT则从计算的角度进行说明。在自组织元表征解释中,高阶大脑网络将低阶网络编码的表征重新编码成元表征,这一过程与意识有关[33, 34]。此外,高阶状态空间理论提出,主观报告(例如,诸如“我感知到了X”之类的陈述)是关于感知内容的生成模型的元认知(高阶)决策[35]。然而,感知现实监测理论(perceptual reality monitoring)假设,当高阶网络将某个一阶表征当做外部世界的可靠反映时,意识感知就会产生[36, 37]。
正如前文所述,HOT专注于解释为什么有些内容是有意识的,而另一些则没有。但下述HOT也不局限于解释这一点——它们也能用来解决有关局部状态的体验特征的问题。一个典型的例子考察了这样一种(有争议的)直觉,即感知体验的内容通常超过“一阶”感官表征中的有效信息,比如周边视觉体验[38,39]。对此,基于HOT的假说认为,周边视觉体验“膨胀”的现象是由一阶状态的高阶表征错误造成的。HOT方法还能解释为什么某些内容是不可能有意识的(它们不能成为适当的元表征状态的目标),以及为什么某些内容必然是有意识的(它们必然伴随有适当的元表征状态)。HOT很少关注意识的全局状态,但他们能够根据(元)表征过程的完整性来解释全局状态之间的区别。
一个特别有趣的问题是HOT能否(如果能,如何)解释各种体验的独特体验特征。为什么欣赏日落的体验特征与头痛的体验特征如此不同?对于这个问题,HOT的回应大体如下:一个状态的体验特征是由相关的元表征状态赋予它的属性决定的。大多数这类解释都集中在视觉体验上[40],但也有解释情绪状态[41]和元认知状态的工作,例如在感知决策过程中,自信的感觉是什么样的[42, 43]。最后,任何完全还原论版的HOT都必须解释为什么各种属性的表征会产生相应的体验,以及神经活动最初如何使这些属性呈现。
HOT主要解释了心智状态具有意识的原因,因此,该方法不提出任何关于意识功能的特定观点。事实上,一些HOT淡化了意识具有独特功能的观点[44]。另一些版本的HOT,通过与信念判断和错误监测相关的元认知过程,来鉴别意识的功能作用[45]。此外,尽管HOT允许有意识的心智状态伴随有意识的元认知判断——例如显式的行为监测或主观信念的报告——但大多数HOT并不要求有意识的感知必然伴随相应的有意识的元认知状态。相反,若元表征想要被意识到,它必须成为合适的元表征状态的对象。
从意识的神经基础角度考虑,对元表征的着重关注导致HOT研究者们着重强调前部皮层区域,特别是前额叶[30],因为这些区域与复杂的认知功能相关。然而,尽管大多数HOT认为前半部分大脑产生意识,但对于具体哪些区域(或过程)是产生意识所必须的,仍有分歧[46]。
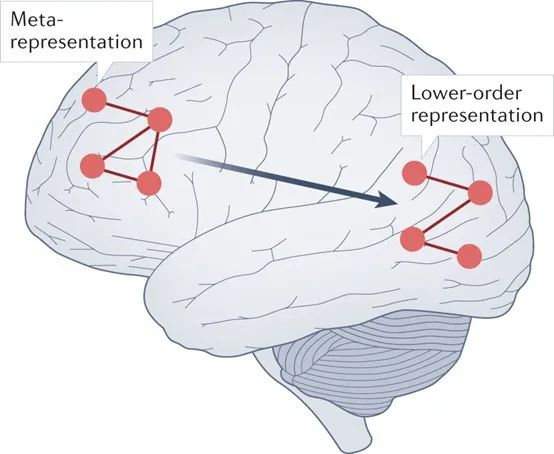
图1:高阶理论。意识的高阶理论(HOT)的核心主张是,心智状态有意识的原因是它成为了特定种类的元表征指向的目标。例如,后部皮层中的视觉信号的低阶表征,在被正确类型的高阶元表征所指向时,将提供有意识的视觉感知。认为前部皮层区域,特别是前额叶区域,参与意识内容的产生的实证研究,是HOT的支持性证据,特别是当有意识和无意识状态都与之匹配时[30,100]。将元认知与前额叶区域联系起来的病变证据,也间接地支持HOT[153]。HOT也受到一些实证研究的挑战,比如某些研究认为前部区域与意识无关[108, 154],它可能只是为了实现主观报告和执行控制而存在[6]。
2. 全局工作空间理论
全局工作空间理论(GWT)起源于人工智能中的“黑板”架构,黑板是一种中央资源库,各种专用处理器通过它共享和接收信息。第一个GWT[47]是在认知层面上构建的,它提出,有意识的心智状态是那些“全局可用”的状态,能被各种认知过程利用,包括注意力、评估、记忆和口头报告。GWT的核心主张是,信息能够被各式各样的作为信息消费者的认知系统所利用,这种可利用性构成了意识体验(图 2)。
这一基本主张后来发展成了神经理论——通常被称为“全局神经元工作空间理论”——根据该理论,当感觉信息在神经元工作空间内“广播”时,感觉信息可以获得意识,该工作空间在解剖学上广泛存在于高级联合皮质内,特别是(但也不只是)前额叶[48, 49]。对全局工作空间的访问需要通过非线性网络“点火”实现,其中的循环处理过程放大并维持神经元表征[50]。与元表征相比,对点火和广播的强调是GWT与HOT方法的差异之一。
与HOT类似,GWT关注意识产生的原因,很少关注不同意识体验之间的差异(除了[51]等)。回到双目竞争实验,GWT旨在解释为什么在特定时刻,对应于“房子”的心智状态有意识(而对应于“人脸”的心智状态没有意识),但并不直接解释看到房子和看到人脸的主观体验差异。
GWT在意识体验的特征问题上相对沉默,但与此同时更关注意识的功能属性。事实上,GWT经常作为“意识通达”[49]的明确解释;也就是说,它能解释为什么某些表征可以(而其它表征不能)被各式各样的作为信息消费者的认知系统灵活使用。GWT赋予意识的核心功能是,意识状态能够以灵活的、语境相关的方式指导认知和行为。GWT还清楚地说明了意识与其它认知过程的关系,例如注意力和工作记忆。注意力对特定信号进行选择和放大,使它们进入工作空间(因此是有意识的);而意识和工作记忆也密切相关,因为被注意到的工作记忆内容是有意识的,它们通过全局工作空间进行广播[49]。
GWT根据工作空间功能完整性的变化来解释意识的全局状态的变化。在神经活动上,意识的全局性丧失有两个方面的反映。一方面是位于额叶与顶叶的,被认为是全局工作空间中的“枢纽”节点的区域,它们的功能连接或动态连接受损[52]。另一方面是功能连接越来越倾向于直接反映潜在的结构连接模式[53-55]。
GWT引出的一个重要问题是,工作空间需要具备哪些条件才能具有“全局”性质[25, 56]。是工作空间所广播的消费者认知系统的数量(和类型)更重要?还是工作空间内发生的广播类型更重要?还是这两个因素都与“全局工作空间”有关?如果我们想知道 GWT能对婴儿、脑损伤个体、接受过裂脑手术的人、非人类动物和人工智能系统的意识做出什么预测,就需要回答这些问题。
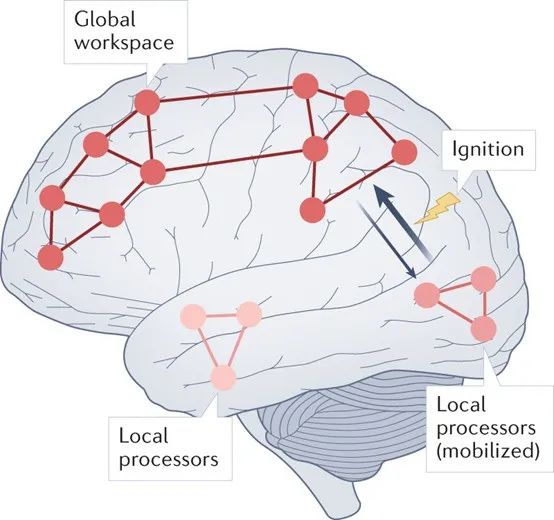
图2:全局工作空间理论。GWT的核心主张是,心智状态在全局工作空间内广播时具有意识,额叶与顶叶区域的神经网络在全局工作空间中扮演着中心枢纽般的角色。当局域处理系统(例如感觉区域)被点火时,其中的活动会暂时性地具有对工作空间的“可移动性”[155]。对GWT的实验证据支持包括,将意识与点火的神经元特征信号,或长程信息共享的神经元特征信号相关联的研究[48, 49, 53, 101]。在外界刺激发生约200-300ms之后,对应于有意识状态或无意识状态的试次,前部皮层区域的神经活动具有差异,这种差异被认为是“点火”的神经元特征信号。这类实验包括“无报告范式”实验[111] (另见[156])。这种研究最近已推广到解码领域。例如,刺激后约300ms的神经活动模式能被用来预测主观报告,而且这种方法可在不同感知模态间泛化[119]。有意识或无意识的内容的长程信息传递特征已经使用一系列方法进行了鉴定[49, 102]。与高阶理论一样,GWT受到前部区域可能参与行为报告而不是意识本身的证据的挑战。
3. 整合信息论
整合信息论(IIT)的出发点与HOT或GWT非常不同——它提出了一种数学方法来描述现象学的特征。该理论先提出了关于意识体验的现象学特征的公理(即,不证自明的、适用于所有可能形式的意识的属性),然后从这些公理推导出任何意识的物理底层所必须满足的属性。接着,IIT提出,将这些属性实例化的物理系统,必然也将意识实例化[57-60](见图3)。具体来说,IIT提出,应该将意识理解为“因果力量”(cause-effect power),与物理系统产生的整合信息(integrated information)的不可化约的最大值相关。反过来,整合信息与信息理论上的数量Φ有关,它衡量的是——广义上——一个系统作为一个整体(而不是独立考虑的各部分),产生多少信息。在IIT中,意识是一个系统的内在的、基本的属性,它既由组成该系统的因果机制的性质决定,也由这些机制的状态决定[60]。
与HOT和GWT相反,IIT主要将意识和后部皮层区域(所谓的“后部热区”,包括顶叶、颞叶和枕叶区域)联系起来。部分原因是,据说,这些区域表现出的神经解剖学特性,很适合产生高层次的整合信息[59]。GWT和HOT(在功能层面)将意识与皮层信息处理的各个方面联系起来。与它们不同,IIT并不是指“信息处理”本身,而是将意识与一个系统的内在因果结构(即一个系统影响自身的因果能力)的属性联系起来。根据IIT,任何能产生最大值非零的(不可化约的)整合信息的系统,至少在某种程度上,都是有意识的。因此,IIT似乎意味着,有意识的非生物系统已经存在[61]。
IIT是相当全面的理论,对意识的全局状态和局部状态都提供了说明 [59](见图3)。全局状态与系统产生的不可化约的整合信息量有关,用Φ来衡量。因此,IIT赞成用单维概念来把握全局状态,因为它把一个有机体的意识水平等同于它的Φ值。而局部状态的体验特征,可以用“概念结构”(conceptual structures)来理解,IIT将其视为高维空间中的“形状”,由系统的机械性因果结构所规定。这些形状产生了(或者说等同于)特定种类的现象特征。例如,视觉体验的空间性质,与早期视觉皮层中的网状机制所规定的因果结构有关[62]。意识的全局统一性是从整合信息的整合角度来解释的——整体产生的信息超过了各个部分产生的信息。最后,当且仅当内容被纳入一个因果“复合体”中,内容才是有意识的(否则则是无意识的)。这儿的复合体是物理系统的一个子集,支撑了最大的不可化约的整合信息。
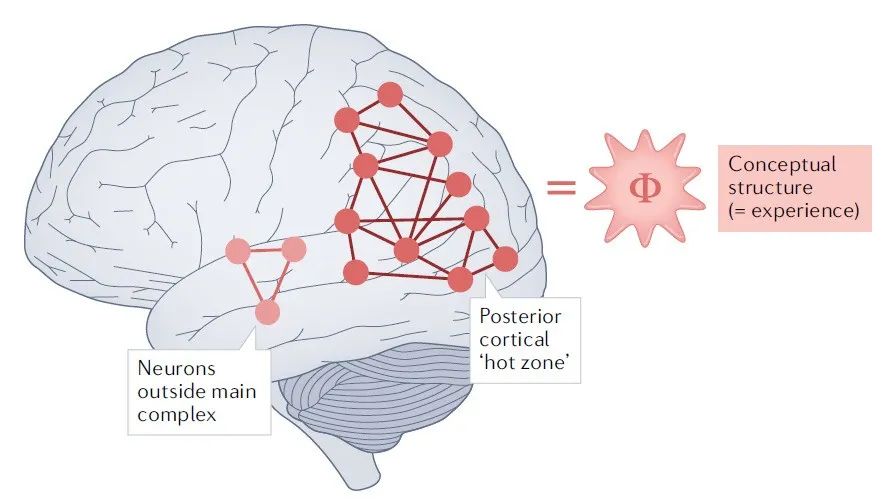
图3:整合信息论。整合信息论(IIT)的核心主张是,意识与物理系统的因果结构是相同的,规定了不可化约的整合信息的最大值。意识的内容与因果结构的形式有关,而意识的水平则与因果结构的不可化约性有关,可用数字Φ来衡量[59]。在解剖学上,IIT与后部皮层的“热区”有关。要对这一核心主张进行实证评估很有挑战性,主要是因为,对Φ的测量是不可行的(除了对某些简单的模型系统)。目前,人们已经开发了各种Φ的替代指标[157],有的指标看起来很有前景。其中,最令人瞩目的是扰动复杂指数(perturbational complexity index,PCI),它测量的是大脑对经颅磁刺激的反应的算法复杂度(Lempel-Ziv)[158]。重要的是,PCI,在跟踪有神经系统损伤的病人的全局意识状态方面,具有诊断和预后价值[158]。然而,PCI并不等同于Φ,而且PCI值与全局意识状态之间的相关性与其它意识理论并不矛盾。一些心理物理学研究间接支持了IIT,证据表明,视觉皮层内侧向连接强度的局部变化,可以改变视觉空间的结构[116],而全局状态的变化,则与后部皮层区域功能多样性和整合能力的降低有关[159]。不过,有证据表明,前部皮层区域的活动是知觉意识所必须的,这将对IIT构成挑战。
回到双目竞争的例子,IIT解释了为什么受试者报告说看到了一栋房子(而不是一张脸)。IIT所主张的假说是,受试者的报告所依据的情结,与对应于内容“房子”(而不是内容“脸”)的概念结构有关。而且,IIT用对应概念结构的“形状”,来解释看到房子和看到脸这两种体验的区别。
虽然IIT对意识的各个方面提供了比大多数ToC更全面的处理,但它对意识和心智的其它方面(如注意力、学习和记忆)的关系说得比较少,而且也没有关注意识的具身性(embodiment)和环境嵌入性(environmental embeddedness)(后两者也是HOT和GWT的挑战)。尽管如此,IIT理论家们已经开始解决其中的一些挑战。例如,他们开发了“匹配复杂度”的测量方法,可以跟踪智能体和其环境之间的共享信息;制定了基于智能体的模型,并发现,能够有效地与周围环境接触的智能体,会表现出更多的整合信息[62-64]。
4. 复馈/预测处理理论
最后,我们考虑两种理解意识的整体路径,它们强调自上而下的信号在塑造和促成意识知觉方面的重要性。第一类——复馈理论(re-entry theories)——本身就是一种ToC,将有意识的知觉与自上而下的(循环的、复馈的)信号联系起来[65, 66]。第二类——预测处理理论(predictive processing theories)——本身不是ToC,但是是对大脑(和身体)功能的更一般的描述,可以用来解释和预测意识的属性[67]。
复馈理论的动机是,神经生理学揭示了自上而下信号对有意识的知觉(通常是视觉)的重要性(例子见参考文献68-70)。在一个著名的复馈理论——局部循环理论(local recurrency theory)中,Lamme认为,在知觉皮层内的局部循环或复馈,足以产生意识(前提是其它有利因素具有完整性,比如脑干的唤醒)。但可能需要顶叶和额叶区域来对知觉体验的内容进行报告,或利用它们进行推理和决策[65,71](图4)。
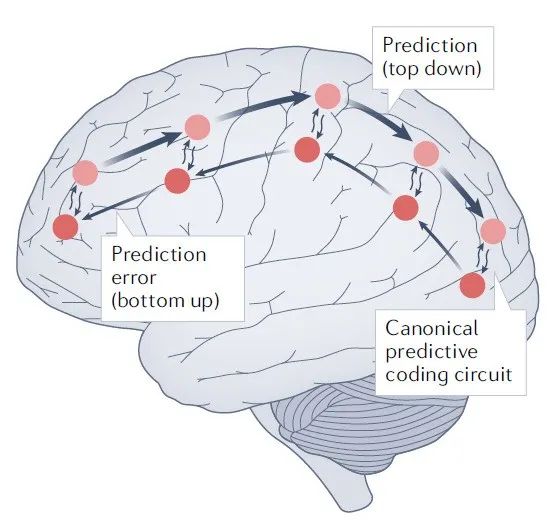
图4:复馈/预测处理理论。复馈理论和预测处理的核心主张是,有意识的心理状态,与自上而下的信号传递(复馈;粗箭头)有关;对于预测处理来说,这些信号传递的,是对感觉信号原因的预测(细箭头表示自下而上的预测误差);因此,对预测误差的不断优化,近似于贝叶斯推理。在大多数预测处理理论中,意识的内容是由自上而下的预测内容所指定的。支持这些理论的证据,来自于一些将自上而下的信号与知觉体验联系起来的研究[68-70, 160]。大量的研究进一步支持了预测处理理论,证据表明,预期(expectation)塑造了意识知觉的内容与速度[161-163],其中,有一些研究将其与自上而下的信号传递直接联系起来[160]。不过,如果有证据表明,自上而下的信号传递或预测处理,会在没有意识的情况下发生,或者这些过程的变化不影响意识状态,那么这些理论就会受到挑战。
广义上讲,预测处理理论有两个动机。一个动机是把感知问题看做是对感觉信号的原因的推断问题[72,73]。另一个动机——以自由能原理(free energy principle)[74]为例——强调在控制和调节方面的基本约束,这些约束适用于所有随着时间推移,能保持其组织的系统[75-77](但也参见文献78)。两者都导致了这样一个概念:通过(通常是自上而下的)知觉预测和(通常是自下而上的)预测误差的相互交流[80](不过参见[81]),大脑实施了一个近似于贝叶斯推理的“预测误差最小化过程”[79]。预测处理的一些表达方式,如主动推理(active inference),增加了一个概念:最小化感官预测误差,不仅可以通过更新预测,还可以通过执行行动,从而控制预测[82,83]。
虽然预测处理理论不是作为ToC产生的,但有人认为,它们可以在神经机制和现象学属性之间,提供系统性的关联[67]。这里的“系统性”指的是在理论的指导下,具有解释力,而不是像经典NCC方法那样,仅仅提供经验性的相关关系。从这个角度看,预测处理理论满足了我们前面概述的ToC的许多要求。不过,最好把它认为是意识科学的理论,而不是意识理论本身,因为就“预测处理和意识到底是什么关系”的问题还有许多争执[84,85]。
预测处理理论,通常以自上而下的知觉预测的内容,来处理局部意识状态[73, 79, 86, 87]。不严格地说,知觉的内容是由大脑对其感官的原因的“最佳猜测”给出的。局部状态的体验特征,是由起作用的知觉预测的性质规定的。例如,视觉中的“客体”现象学,可以由对行动的感觉后果的条件性预测来解释[87,88],而情绪状态的现象学,可以由对内感受的预测在调节有机体的生理状况中的作用来解释[89,90]。情感的例子突出表明,预测处理理论,比起本文讨论的其它理论,更多地涉及了关于有意识自我的问题[13, 77, 91]。
预测处理可以解释有意识和无意识状态之间的区别,取决于在感知推理的过程中,一个心理状态是否是当前“最佳猜测”(或最佳后验)的一部分。在双眼竞争的例子中,预测处理设想了两个相互竞争的知觉假设(最佳猜测),其中一个假设“获胜”时,相应的知觉占据主导地位。此时,来自另一个假设的感觉信号作为预测误差不断积累,最终导致知觉的转变。与此同时,以前占优势的最佳猜测所解释的感觉信号,成为了无法解释的错误预测的来源。就这样循环往复……[92, 93]。(至于房子和脸之间在体验上的差别,如上所述,可以由相应的知觉预测的属性来解释。)在那些强调主动推理的预测处理理论变体中,只有由行为导致的知觉信念变化,才会引起意识内容的变化[76, 94]。(行动可以是明显的,比如眼球的扫动,也可以是隐蔽的,比如注意力焦点的转移。)
预测处理理论一般不涉及意识的整体状态,但这些理论要解释整体状态的区别的话,可以很自然地诉诸相关预测过程的完整性[95],就像HOT理论可以诉诸相关元表征机制的完整性。
至于意识的功能层面,复馈/预测处理理论都对意识和注意力之间的关系进行了明确的区分。在局部循环理论中,注意力为感觉信号提供了一个选择性的增强(正如在GWT中一样),从而使它们到达前额叶和顶叶区域,参与意识的访问[71]。在预测处理中,注意力与“精确度加权”(precision weighting)的过程有关。这个过程调制了感觉信号的估计精度(estimated precision),直观来说,其方式相当于改变这些信息的信噪比或“增益”[74, 96]。而在主动推理中,如前所述,注意力抽样可能是意识内容变化的必要条件[76, 94]。
四、意识理论的评估标准和发展方向
在对立的ToC之间的争论中,研究者呼吁使用的数据的范围很广,所以我们并不奢望在这里就提供一个完整的清单。相反,我们对目前的一些争论进行了有选择的概述,强调了可以用来评估ToC的数据的多样性。(图1-4的图例中,描述了其它一些通常用来支持各个ToC的实验数据。)
有一个点需要事先说明:认识到理论评价的整体性是很重要的。理论不会被一个单一的发现所证实,通常也不会被一个单一的实验所打败。相反,对理论的确认通常是一个渐进的过程。在这个过程中,一个理论通过提供对目标现象的解释战胜它的对手;这个解释需要合理解释广泛的数据,并能够和邻近领域的成功理论相结合[97-99]。
一个明显的制约ToC的因素是意识的结构。尽管我们在讨论ToC时,已经讨论了许多结构性特征,但要对比各种ToC,一个特别有用的结构性特征,是意识的统一性——一个智能体,在某一时刻的各种体验,似乎总是作为一个统一的复杂体验的组成部分而发生,这个体验就完全是对“成为这个智能体是什么感觉”的回答[25]。不同的ToC,对意识的统一性采取非常不同的态度。IIT相当强调意识的统一性,它不仅假定意识始终是统一的,而且还呼吁“意识必然是统一的”这一主张,从而使意识与不可化约的整合信息(最大值)得以相互关联。尽管GWT没有像IIT那么强调意识的统一性,但意识与功能整合的工作空间的广播的关联表明,这类理论也可能提供一个关于意识统一性的可信的解释。其它ToC,例如HOT和复馈/预测处理理论,与意识的统一性有着更矛盾的关系,要么只是假装解释这一属性,要么完全忽略它。各个ToC对意识的统一性的态度的不同,至少部分是因为更根本的分歧——意识是否(必然)统一。尽管意识的统一性有望为ToC提供一个重要的约束,但为了兑现这一前景,我们需要在“意识(必然)统一”这方面有一个更好地说明。
第二个制约因素来源于神经数据。例如,人们普遍认为,小脑对意识来说,既不必要,也不充分。一个ToC应该解释这一事实,并解释为什么小脑不与意识有关。一些ToC很容易提供这样的解释——例如,IIT认为,小脑与意识无关,因为它的结构不适合产生高水平的整合信息[59]。但这种解释,只有在它比其它理论可能提供的解释更可信的情况下,才会对某一理论提供具体的支持,而这一条件是否满足,目前还是一个开放的问题。例如,主张HOT的人可以认为,小脑缺乏支持相关类型的元表征的能力;主张GWT的人可以提出,小脑没有实现全局工作空间的理由;而复馈/预测处理理论家可以指出,小脑中缺乏丰富的循环信号[65]。
尽管人们普遍认为,ToC应该解释为什么小脑与意识无关,但从ToC的角度来看,还有其它类型的神经数据更具有争议性。一个重要的例子是关于前额叶(“大脑前部”)过程在意识中的作用的讨论。
许多神经影像学研究,根据额叶的区域活动[100, 101]和额叶与其它区域的功能连接,利用各种实验范式,发现前额叶会参与有意识(相对于无意识)的感知[48]。少数对灵长类动物的研究也发现,在双目竞争、连续闪光的抑制和视觉刺激的快速连续呈现的过程中,有意识的内容可以从前额叶的活动模式中解码出来[103-105]。参考文献[106]则提供了一个更复杂的图景:在一个物体识别任务中,内容相关的信息可以从大范围的激活和失活的皮层区域中解码出来。而病变的证据和脑刺激的证据也被用来论证,前额叶的活动与意识有重要的关系(见参考文献30)。
倡导HOT和GWT的人用这些发现来支持他们的理论,而不是与其竞争的理论。作为回应,IIT和复馈理论的倡导者认为,观察到的前额叶活动是意识的一个(非必要的)结果,而不是与有意识的知觉本身有关[107, 108](但见参考文献109)。那些为这种“大脑后部”观点辩护的人认为,后部皮层过程——涉及楔前区和一部分的知觉皮层和顶叶皮层——对知觉体验来说就已经足够了,而“大脑前部”过程是不必要的。这种说法得到了所谓“无报告”研究的支持。这些研究通常会发现,当受试者不提供关于他们知觉的明确报告时,前额叶的参与会减少[6, 110](但见参考文献111)。“大脑后部”的倡导者也利用积极的证据,来证明后部活动和意识之间的紧密联系。例如,一项新研究使用一种连续唤醒的范式来探测睡眠期间的意识内容,发现后部皮层区域的活动可以预测一个人是否会报告梦的经历,包括在快速眼动和非快速眼动睡眠阶段[112](但见参考文献113)。最后,解码研究中的“大脑前部”解释是否成立还有待商榷。这是因为,表明意识内容可以从一个特定的区域“读出”,并不能确定大脑本身正以一种构成相关元表征或全局广播的方式,从该区域“读出”这些内容。
尽管“大脑前部”和“大脑后部”的争论在某些方面确实涉及神经生物学证据——例如,对于前额叶皮层的边界具体在哪里,存在不同的意见[107, 109],但其核心还是对意识和认知访问(cognitive access)之间关系的分歧:将可用于口头报告的内容和对行为的直接控制作为意识的替代是否合理?对意识的神经基础的研究,是不是应该在“意识和认知访问究竟如何关联”方面保持中立[114](亦可参见方框3)?关于这个问题的争论,反应在不同的ToC对认知访问的态度上。GWT把认知摆在核心位置,不仅表明意识的内容总是可以被认知访问,而且表明支撑认知访问的过程本身(即点火和全局广播)就是意识体验的基础(关于这一观点的最近细微差别,见参考文献111)。其它理论,如IIT和局部循环理论,否认意识和认知访问之间的密切关系,认为心智状态在不直接控制思想和行动的情况下也可以有意识,也认为心智状态原则上即使不具有意识,也可以直接控制思想和行动。尽管HOT并不执着于意识和认知访问之间的任何特定关系,但在实践中,这类理论的倡导者通常假定,意识的内容将被认知访问(例如参考文献46),而不是反过来。
要评估相互竞争的ToC,也许最有力的证据是新的预测。科学史上许多最重要的事件都涉及对新预测的确认[115]。例如,广义相对论既预测了水星近日点的进动,又预测了掠过太阳表面星光的偏转方式,从这些事实中获得了强有力的支持。如果一个ToC做出了新预测并被证实,那么它就会得到强有力的支持,特别是与那些未能做出相关预测,或做出不同的、不相容预测的理论相比较时。
当代ToC所做的许多新的预测都难以测试。例如,复馈理论和IIT都预测,后部皮层活动可以在没有前部区域贡献的情况下支持意识体验,但目前我们缺乏可靠的方法来验证这种说法,因为验证依赖于主观报告(或者至少是某种形式的执行控制),而这又需要前部皮层活动。更引人注目的是,IIT预测,意识广泛分布于整个自然界,包括许多非生物系统,甚至可能出现在简单的系统(如光电二极管和单原子)中。有趣的是,不出现在严格意义上的前馈神经网络中[61]。这一预测与大家广泛认同的对意识分布的假设是相悖的,但由于缺乏有力的方法来检测此类系统是否存在意识,因此无法对其进行合理的评估(方框3)。
方框3
测量问题
为了检验意识理论,我们需要可靠地检测意识的存在和不存在,目前,实验者通常依靠受试者的内省能力,直接或间接地来确定其意识状态。然而,这种方法是有问题的,因为内省的可靠性值得怀疑,而且有许多生物体或系统(例如婴儿、有脑损伤的个体和非人类的动物)可能有意识,但无法产生内省报告。因此,人们迫切需要确定不依靠内省的“标记”或意识的“特征”。近年来,人们提出了许多这样的指标,提议其中一些——如扰动复杂度指数(perturbational complexity index,PCI)[158]——作为意识本身的标记,而提议其它指标——如视动眼球震颤反应(optokinetic nystagmus response)[174]或神经动力学中的独特分叉(distinctive bifurcations in neural dynamics)[111]——作为特定种类的意识内容的标记。人们已经卓有成效地将前者应用于评估脑损伤患者的整体意识状态[175],而将后者用于意识内容的“无报告”研究,即没有明确的行为报告的研究[6]。然而,无论重点是什么,任何提出来的意识指标,都必须得到验证:我们需要确认它既灵敏又明确。尽管基于内省的验证方法存在前面所述的问题,但基于理论的方法也存在问题。由于ToC本身就有争议,因此,这些基于理论的指标也难以提供意识的客观标记物所需要的那种跨越不同主体的验证方式。因此,解决测量问题似乎需要一种既不完全基于内省,也不完全基于理论考虑的验证方法。文献中包含了一些解决这个问题的提议[114, 176],但没有一个是毫无争议的[177, 178]。
在某些情况下,方法上的进步可能会带来达到可测试程度的新预测。例如,从IIT产生的一个引人注目的预测是,神经结构的变化可能导致意识体验的变化,即使前者并没有引起神经活动的变化[116]。例如,视觉皮层中不活跃的(inactive)神经元可能对视觉体验有贡献,但失活的(inactivated)神经元则不会[59]。IIT产生这一预测是因为,对意识来说,由神经机制规定的因果结构才是重要的。这意味着,如果我们干预神经机制,从而改变因果结构,那么即使相应的神经动力学没有变化,意识也会发生变化。这一预测在没有动力学的情况下(即对于不活跃的神经元)尤其反直觉。这一猜想可能可以使用精确的干预方式(如光遗传学),在知觉决策的动物模型中进行测试[117]。而本文所讨论的其它理论并不容易得出类似的猜想。
评估相互竞争的ToC的一个特别有效的途径是,关注有意识(相对于无意识)过程的时间特征,例如电生理记录中的时间相关电位。一些理论家(例如,见参考文献118)认为,有意识的知觉在刺激呈现后的早期(120-200毫秒)就开始了,并认为有证据表明,知觉意识和早发的、模式特定的、负向的、事件相关的电位(称之为意识负性反应)之间存在着强有力的相关性;他们还质疑了以往讨论的晚发特征的可靠性,如P3b(在刺激开始后约300毫秒观察到的正向的、事件相关的点位)。Dembski及其同事强调的早期负性(early negativity)在视觉和听觉中都有发现,这使得他们认为,存在一种普遍的早发反应,能有力地反映出知觉的意识。这样的证据有利于IIT和局部复馈解释(但见参考文献119,讲了后来研究的意识知觉的跨模态特征)。其他理论家[120, 121]则主张知觉意识的启动时间要晚得多(大约是250-400毫秒)。除了有争议的P3b,晚发的理论似乎是由各种知觉现象引起的,这些现象与这一时间尺度相匹配,包括心理不应期(psychological refractory)、注意瞬脱(attentional blink)和事后预测效应(postdictive effects )。后者能有意义地表明,延迟的线索可以回溯性地触发有意识的知觉[122]。有意识知觉的晚发神经特征的候选者包括长距离的信息共享和分叉活动[49, 111]。支持晚发知觉意识的证据,通常支持高阶和全局工作空间的意识理论。在可预见的未来,关于知觉体验的“早发”和“晚发”说法之间的争论,可能仍然是讨论的中心。请注意,意识处理的时间特征问题,与对持续时间的感知[123]和意识“时刻”的时间特征都是不同的[124, 125],后两者反映的是意识内容方面,应该由ToC来解释。
五、展望
目前,在意识科学中,ToC通常被用作“叙述结构”。尽管它们为神经和行为数据提供了解释,但在设计研究时考虑到理论验证的情况仍然很少[126]。尽管以这种方式运用意识理论并没有什么不妥,但未来的研究进展,将取决于那些能够对ToC进行测试和区分的实验。在这篇综述中,我们确认了三个需要解决的问题,敦促意识科学中成熟的理论测试制度的发展。
首先,ToC的发展需要精度,因为那些构造模糊而不精确的理论,只能产生模糊和不精确的预测。例如,HOT与复馈/预测处理理论需要明确(特定方面的)意识所独有的元表征和复馈或预测性过程的种类;IIT需要精确地说明它对意识的功能特征意味着什么,以及环境和意识的具身性的影响;GWT需要提供一个原则性说明,哪些工作空间,在相关意义上,有资格称为“全局”。
一个有前景的方法是,使用计算模型,为可能以相对抽象或概念性的术语表述的ToC带来机制上的具体性。除了为产生精细预测奠定基础外,这种模型还可以提供一种共同的语言。用这种语言,可以比较相互竞争的ToC的相对优点,这对于比较理论出发点不一致的ToC特别有用。例如,计算模型可以揭示HOT和复馈/预测处理理论中自上而下信号的共同原则,同时澄清元表征(例如,见参考文献35)和全局广播(例如,见参考文献127, 128)之间的区别,将HOT和GWT分开[129]。计算模型的发展,也可能使得ToC之间的对比从(潜在的分布式)过程的角度来重构,而不是从广泛的神经解剖学区域的角度来重构(就像目前流行的那样,例如,“大脑前部”和“大脑后部”理论家之间的辩论[111])。计算方法的一个关键挑战,是开发出不仅能解释意识的功能特征,而且能解释其现象学属性的模型。这一挑战可以用“计算(神经)现象学”这个广义标签来描述(例子见参考文献37,130)。这就带来了额外的挑战,即如何用现象学的数据来验证计算模型,或者在计算模型之间进行区分(例子见参考文献131)。这一挑战,至少可以部分地通过在适当的现象学颗粒度上收集主观报告来应对(方框3)。
除了变得更加具体,ToC还需要变得更加全面。在大多数情况下,ToC倾向于关注特定类型的局部状态(感知体验,重点是视觉)、特定类型的全局状态(普通的清醒意识)和特定类型的有意识的生物(成年人类)。尽管理论家们对于倾向于关注有限的意识状态和生物有很好的理由——实验的可及性,但一个全面的ToC必须公正地对待丰富多样的意识。就局部状态而言,ToC必须超越感知,对情感、时间性、意志和思想等状态进行解释。在全局状态方面,ToC必须超越普通的清醒状态,对做梦、冥想、意识紊乱和迷幻状态等独特的意识模式进行解释。关于有意识的生物的类型,ToC必须超越成年人的体验,解决人类婴儿、非人类动物甚至人工系统的意识问题。尽管关注点有限的ToC并无不妥,但能对意识提供全面说明的理论具有明显的优势,尤其是当它们能够识别并解释意识的不同方面之间的联系时。
第三个需要解决的问题是测量问题:找到值得信赖的意识测量方法[132]。解决这个问题至关重要,因为,除非我们有能力验证其预测,否则一个详细而全面的ToC也不会有多大用处。区分测量问题的两个(密切相关的)版本是有用的。第一个是对意识内容的检测。在这里,主要的挑战,是找出方法能区分有意识和无意识心理状态,而且不对意识的功能特征做出有争议的假设(如,有意识的内容必须是可报告的,或是可用于高级认知控制的)[114, 133, 134]。测量问题的另一个版本关注的不是内容,而是生物。问题包括,如何确定意识在动物界的分布[135];类大脑器官[136]或人工智能系统[135-137]是否有意识;意识何时在个体发育中首次出现[138];以及意识怎样会在脑外伤的情况下还得以保留[139]。这里的挑战依然在于,要开发能测量意识的方法,又要避免对意识的功能特征的争议性假设(方框3)。
当然,意识领域的研究人员已经在不同程度上处理了上述的挑战。这些尝试目前得到了一些举措的补充,如对抗性合作模式。该模式正在鼓励ToC的支持者设计实验,来区分不同的ToC[17]。在科学上,意识仍然有争议,但我们有理由认为,ToC的迭代发展、测试和比较,将使我们更深地理解这个最深刻的奥妙。
核心概念列表
意识的神经相关物(Neural correlates of consciousness)
共同足以构成某种意识状态的最小神经事件集
解释鸿沟直觉(Explanatory gap intuitions)
认为不可能用物理的、机械的术语给予意识完全令人满意的解释的直觉。
对抗式合作(Adversarial collaborations)
一类研究项目。其中,不同理论的支持者共同设计一个实验来区分他们喜欢的理论,而且,对实验结果如何更有利于哪一种理论,他们需要事先达成一致。
全局状态(Global states)
与生物体的整体意识状态有关,通常与唤醒和行为反应有关,并与意识的“水平”有关。
局部状态(Local states)
与特定的有意识的心智状态有关,例如有意识的知觉、情感或思想。局部状态也经常被称为有意识的内容。
双目竞争(Binocular rivalry)
向两只眼睛分别呈现不同的图像,有意识的感知将在两个图像之间交替,这种现象叫双目竞争。
现象特征(Phenomenal character)
局部状态的主观体验特质,比如看到红色时感受到“红色的感觉”或牙痛时感受到疼痛——有时也称为感受性质(qualia)。
元表征(Meta-representation)
以另一种心智表征为目标的心智表征。
无报告范式(No-report paradigms)
被试不提供主观(口头、行为)报告的行为实验。
Φ
指的是一个系统所规定的,不可由它的部件所规定的信息量所化约的信息量。Φ有许多变体,每一种都有着不同的计算方法,做出不同的假设。
后部热区(Posterior hot zone)
靠近大脑皮层后部的一系列脑区,包括顶叶、颞叶和枕叶区,以及楔前区等区域。
复合体(Complex)
在整合信息论(IIT)中,支撑着最大的不可化约的整合信息的一个物理系统的一个子集。
意识的统一性(Unity of consciousness)
指的是下列事实:一个智能体在某一时期的各种体验,似乎总是作为单个复杂体验的组成部分而发生。
认知访问(Cognitive access)
指的是一种功能属性,即一种心理状态可以接触到一系列认知过程,通常包括语言和/或行为报告。
内感受预测(Interoceptive predictions)
指的是对源自身体内部的感觉信号的原因的预测(内感受指的“从内部”对身体的感知)。
计算(神经)现象学(Computational (neuro) phenomenology)
指的是使用计算模型从(神经)机制的角度来解释意识状态的现象学特征。
测量问题(The measurement problem)
指的是确认某一心智状态是否是有意识的的问题,或确认某一有机体或系统,是否是,或者是否有能力是有意识的的问题。
类大脑器官(Cerebral organoids)
实验室生长的神经结构,它能自我组织成具有细胞和网络特征的系统,类似于发育中的人脑。
参考文献
1. Crick, F. & Koch, C. Towards a neurobiological theory of consciousness. Semin. Neurosci. 2, 263–275 (1990).
2. Metzinger, T. (ed.) Neural Correlates of Consciousness: Empirical and Conceptual Questions (MIT Press, 2000).
3. Koch, C., Massimini, M., Boly, M. & Tononi, G. Neural correlates of consciousness: progress and problems. Nat. Rev. Neurosci. 17, 307–321 (2016).
4. de Graaf, T. A., Hsieh, P. J. & Sack, A. T. The ‘correlates’ in neural correlates of consciousness. Neurosci. Biobehav. Rev. 36, 191–197 (2012). 5. Aru, J., Bachmann, T., Singer, W. & Melloni, L. Distilling the neural correlates of consciousness. Neurosci. Biobehav. Rev. 36, 737–746 (2012).
6. Tsuchiya, N., Wilke, M., Frassle, S. & Lamme, V. A. No-report paradigms: extracting the true neural correlates of consciousness. Trends Cogn. Sci. 19, 757–770 (2015).
7. Klein, C., Hohwy, J. & Bayne, T. Explanation in the science of consciousness: from the neural correlates of consciousness (NCCs) to the difference-makers of consciousness (DMCs). Philos. Mind Sci. https://doi.org/10.33735/phimisci.2020.II.60 (2020).
8. Michel, M. et al. Opportunities and challenges for a maturing science of consciousness. Nat. Hum. Behav. 3, 104–107 (2019).
9. Seth, A. K. Consciousness: the last 50 years (and the next). Brain Neurosci. Adv. 2, 2398212818816019 (2018).
10. Seth, A. K. Explanatory correlates of consciousness: theoretical and computational challenges. Cogn. Comput. 1, 50–63 (2009).
11. Searle, J. The Rediscovery of the Mind (MIT Press, 1992).
12. Varela, F. J. Neurophenomenology: a methodological remedy for the hard problem. J. Conscious. Stud. 3, 330–350 (1996).
13. Seth, A. K. Being You: A New Science of Consciousness (Faber & Faber, 2021).
14. Dennett, D. C. Welcome to strong illusionism. J. Conscious. Stud. 26, 48–58 (2019).
15. Frankish, K. Illusionism as a Theory of Consciousness (Imprint Academic, 2017).
16. Wiese, W. The science of consciousness does not need another theory, it needs a minimal unifying model. Neurosci. Conscious. 2020, niaa013 (2020).
17. Melloni, L., Mudrik, L., Pitts, M. & Koch, C. Making the hard problem of consciousness easier. Science 372, 911–912 (2021).
This work sets out how an adversarial collaboration is planning to arbitrate between integrated information and global workspace ToCs.
18. Hameroff, S. & Penrose, R. Consciousness in the universe: a review of the 'Orch OR' theory. Phys. Life Rev. 11, 39–78 (2014).
19. Chalmers, D. J. & McQueen, K. in Quantum Mechanics and Consciousness (ed Gao, S.) (Oxford Univ. Press, 2022).
20. Nagel, T. What is it like to be a bat? Philos. Rev. 83, 435–450 (1974).
21. Bayne, T., Hohwy, J. & Owen, A. M. Are there levels of consciousness? Trends Cogn. Sci. 20, 405–413 (2016).
This work challenges the common unidimensional notion of 'level of consciousness', outlining an alternative, richer, multidimensional account.
22. Metzinger, T. Being No-One (MIT Press, 2003).
23. Damasio, A. Self Comes To Mind: Constructing the Conscious Brain (William Heinemann, 2010).
24. Park, H. D. & Tallon-Baudry, C. The neural subjective frame: from bodily signals to perceptual consciousness. Philos. Trans. R. Soc. Lond. B Biol. Sci. 369, 20130208 (2014).
25. Bayne, T. The Unity of Consciousness (Oxford Univ. Press, 2010).
26. Bayne, T. & Chalmers, D. J. in The Unity of Consciousness: Binding, Integration, and Dissociation (ed Cleeremans, A.) 23–58 (Oxford Univ. Press, 2003).
27. Cummins, R. Functional analysis. J. Philos. 72, 741–765 (1975).
28. Blake, R., Brascamp, J. & Heeger, D. J. Can binocular rivalry reveal neural correlates of consciousness? Philos. Trans. R. Soc. Lond. B Biol. Sci. 369, 20130211 (2014).
29. Signorelli, C. M., Szczotka, J. & Prentner, R. Explanatory profiles of models of consciousness — towards a systematic classification. Neurosci. Conscious. 2021, niab021 (2021).
30. Lau, H. & Rosenthal, D. Empirical support for higherorder theories of conscious awareness. Trends Cogn. Sci. 15, 365–373 (2011).
This work presents a summary of empirical evidence favouring higher-order ToCs.
31. Rosenthal, D. Consciousness and Mind (Clarendon, 2005).
32. Brown, R. The HOROR theory of phenomenal consciousness. Philos. Stud. 172, 1783–1794 (2015).
33. Cleeremans, A. Consciousness: the radical plasticity thesis. Prog. Brain Res. 168, 19–33 (2008).
34. Cleeremans, A. et al. Learning to be conscious. Trends Cogn. Sci. 24, 112–123 (2020).
35. Fleming, S. M. Awareness as inference in a higherorder state space. Neurosci. onscious. 2020, niz020 (2020).
36. Lau, H. Consciousness, metacognition, and perceptual reality monitoring. Preprint at ArXiv https://doi.org/10.31234/osf.io/ckbyf (2020).
37. Gershman, S. J. The generative adversarial brain. Front. Artif. Intell. https://doi.org/10.3389/frai.2019.00018 (2019).
38. Cohen, M. A., Dennett, D. C. & Kanwisher, N. What is the bandwidth of perceptual experience? Trends Cogn. Sci. 20, 324–335 (2016).
39. Haun, A. M., Tononi, G., Koch, C. & Tsuchiya, N. Are we underestimating the richness of visual experiences? Neurosci. Conscious. 3, 1–4 (2017).
40. Odegaard, B., Chang, M. Y., Lau, H. & Cheung, S. H. Inflation versus filling-in: why we feel we see more than we actually do in peripheral vision. Philos. Trans. R. Soc. Lond. B Biol. Sci. https://doi.org/10.1098/rstb.2017.0345 (2018).
41. LeDoux, J. E. & Brown, R. A higher-order theory of emotional consciousness. Proc. Natl Acad. Sci. USA 114, E2016–E2025 (2017).
42. Morrison, J. Perceptual confidence. Anal. Philos. 78, 99–147 (2016).
43. Peters, M. A. K. Towards characterizing the canonical computations generating phenomenal experience. Preprint at PsyArXiv https://doi.org/10.31234/osf.io/bqfr6 (2021).
44. Rosenthal, D. Consciousness and its function. Neuropsychologia 46, 829–840 (2008).
45. Charles, L., Van Opstal, F., Marti, S. & Dehaene, S. Distinct brain mechanisms for conscious versus subliminal error detection. Neuroimage 73, 80–94 (2013).
46. Brown, R., Lau, H. & LeDoux, J. E. Understanding the higher-order approach to consciousness. Trends Cogn. Sci. 23, 754–768 (2019).
47. Baars, B. J. A Cognitive Theory of Consciousness (Cambridge Univ. Press, 1988).
48. Dehaene, S. & Changeux, J. P. Experimental and theoretical approaches to conscious processing. Neuron 70, 200–227 (2011).
49. Mashour, G. A., Roelfsema, P., Changeux, J. P. & Dehaene, S. Conscious processing and the global neuronal workspace hypothesis. Neuron 105, 776–798 (2020).
This work presents a summary of the neuronal GWT and its supporting evidence.
50. Dehaene, S., Sergent, C. & Changeux, J. P. A neuronal network model linking subjective reports and objective physiological data during conscious perception. Proc. Natl Acad. Sci. USA 100, 8520–8525 (2003).
51. Naccache, L. Why and how access consciousness can account for phenomenal consciousness. Philos. Trans. R. Soc. Lond. B Biol. Sci. https://doi.org/10.1098/rstb.2017.0357 (2018).
52. Mashour, G. A. Cognitive unbinding: a neuroscientific paradigm of general anesthesia and related states of unconsciousness. Neurosci. Biobehav. Rev. 37, 2751–2759 (2013).
53. Demertzi, A. et al. Human consciousness is supported by dynamic complex patterns of brain signal coordination. Sci. Adv. 5, eaat7603 (2019).
This large empirical study of functional connectivity patterns across different global states of consciousness focuses on how these patterns relate to underlying structural connectivity.
54. Barttfeld, P. et al. Signature of consciousness in the dynamics of resting-state brain activity. Proc. Natl Acad. Sci. USA 112, 887–892 (2015).
55. Uhrig, L. et al. Resting-state dynamics as a cortical signature of anesthesia in monkeys. Anesthesiology 129, 942–958 (2018).
56. Carruthers, P. Human and Animal Minds: The Consciousness Questions Laid to Rest (Oxford Univ. Press, 2019).
57. Tononi, G. Consciousness as integrated information: a provisional manifesto. Biol. Bull. 215, 216–242 (2008).
58. Tononi, G. Integrated information theory of consciousness: an updated account. Arch. Ital. Biol. 150, 293–329 (2012).
59. Tononi, G., Boly, M., Massimini, M. & Koch, C. Integrated information theory: from consciousness to its physical substrate. Nat. Rev. Neurosci. 17, 450–461 (2016).
This work presents an account of the core claims and concepts of the integrated information ToC.
60. Oizumi, M., Albantakis, L. & Tononi, G. From the phenomenology to the mechanisms of consciousness: integrated information theory 3.0. PLoS Comput. Biol.
10, e1003588 (2014).
61. Tononi, G. & Koch, C. Consciousness: here, there and everywhere? Philos. Trans. R. Soc. Lond. B Biol. Sci. https://doi.org/10.1098/rstb.2014.0167 (2015).
62. Haun, A. M. & Tononi, G. Why does space feel the way it does? Towards a principled account of spatial experienc. Entropy 21, 1160 (2019).
63. Albantakis, L., Hintze, A., Koch, C., Adami, C. & Tononi, G. Evolution of integrated causal structures in animats exposed to environments of increasing complexity. PLoS Comput. Biol. 10, e1003966 (2014).
64. Marshall, W., Gomez- Ramirez, J. & Tononi, G. Integrated information and state differentiation. Front. Psychol. 7, 926 (2016).
65. Lamme, V. A. Towards a true neural stance on consciousness. Trends Cogn. Sci. 10, 494–501 (2006).
66. Lamme, V. A. & Roelfsema, P. R. The distinct modes of vision offered by feedforward and recurrent processing. Trends Neurosci. 23, 571–579 (2000).
67. Hohwy, J. & Seth, A. K. Predictive processing as a systematic basis for identifying the neural correlates of consciousness. Philos. Mind Sci. 1, 3 (2020).
68. Lamme, V. A., Super, H., Landman, R., Roelfsema, P. R. & Spekreijse, H. The role of primary visual cortex (V1) in visual awareness. Vis. Res. 40, 1507–1521 (2000).
69. Pascual- Leone, A. & Walsh, V. Fast backprojections from the motion to the primary visual area necessary for visual awareness. Science 292, 510–512 (2001).
This early study uses transcranial magnetic stimulation to reveal a role for re- entrant activity in conscious visual perception in humans.
70. Boehler, C. N., Schoenfeld, M. A., Heinze, H. J. & Hopf, J. M. Rapid recurrent processing gates awareness in primary visual cortex. Proc. Natl Acad. Sci. USA 105, 8742–8747 (2008).
71. Lamme, V. A. How neuroscience will change our view on consciousness. Cogn. Neurosci. 1, 204–220 (2010).
72. von Helmholtz, H. Handbuch der Phsyiologischen Optik [German] (Voss, 1867).
73. Clark, A. Whatever next? Predictive brains, situated agents, and the future of cognitive science. Behav. Brain Sci. 36, 181–204 (2013).
This work presents a classic exposition of predictive processing and its relevance for perception, cognition and action.
74. Friston, K. J. The free- energy principle: a unified brain theory? Nat. Rev. Neurosci. 11, 127–138 (2010).
75. Seth, A. K. in Open MIND (eds Windt, J. M. & Metzinger, T.) (MIND Group, 2015).
76. Friston, K. J. Am I self- conscious? (Or does self- organization entail self- consciousness?). Front. Psychol. 9, 579 (2018).
77. Seth, A. K. & Tsakiris, M. Being a beast machine: the somatic basis of selfhood. Trends Cogn. Sci. 22, 969–981 (2018).
78. Bruineberg, J., Dolega, K., Dewhurst, J. & Baltieri, M. The Emperor’s new Markov blankets. Behav. Brain Sci. https://doi.org/10.1017/S0140525X21002351 (2021).
79. Hohwy, J. The Predictive Mind (Oxford Univ. Press, 2013).
80. Rao, R. P. & Ballard, D. H. Predictive coding in the visual cortex: a functional interpretation of some extra- classical receptive- field effects. Nat. Neurosci. 2,
79–87 (1999).
81. Teufel, C. & Fletcher, P. C. Forms of prediction in the nervous system. Nat. Rev. Neurosci. 21, 231–242 (2020).
82. Friston, K. J., Daunizeau, J., Kilner, J. & Kiebel, S. J. Action and behavior: a free- energy formulation. Biol. Cybern. 102, 227–260 (2010).
83. Parr, T. & Friston, K. J. Generalised free energy and active inference. Biol. Cybern. 113, 495–513 (2019).
84. Pennartz, C. M. A. Consciousness, representation, action: the importance of being goal- directed. Trends Cogn. Sci. 22, 137–153 (2018).
85. Williford, K., Bennequin, D., Friston, K. & Rudrauf, D. The projective consciousness model and phenomenal selfhood. Front. Psychol. 9, 2571 (2018).
86. Hohwy, J. New directions in predictive processing. Mind Lang. 35, 209–223 (2020).
87. Seth, A. K. A predictive processing theory of sensorimotor contingencies: explaining the puzzle of perceptual presence and its absence in synesthesia. Cogn. Neurosci. 5, 97–118 (2014).
88. O’Regan, J. K. & Noë, A. A sensorimotor account of vision and visual consciousness. Behav. Brain Sci. 24, 939–973; discussion 973–1031 (2001).
This primary description of the sensorimotor ToC argues that conscious perception is intimately related to action.
89. Seth, A. K. Interoceptive inference, emotion, and the embodied self. Trends Cogn. Sci. 17, 565–573 (2013).
This work presents a theoretical application of predictive processing to interoception and physiological regulation, relating this to experiences of emotion and selfhood.
90. Barrett, L. F. The theory of constructed emotion: an active inference account of interoception and categorization. Soc. Cogn. Affect. Neurosci. 12, 1833 (2017).
91. Solms, M. The hard problem of consciousness and the free energy principle. Front. Psychol. 9, 2714 (2018).
92. Hohwy, J., Roepstorff, A. & Friston, K. Predictive coding explains binocular rivalry: an epistemological review. Cognition 108, 687–701 (2008).
93. Parr, T., Corcoran, A. W., Friston, K. J. & Hohwy, J. Perceptual awareness and active inference. Neurosci. Conscious. 2019, niz012 (2019).
94. Friston, K. J., FitzGerald, T., Rigoli, F., Schwartenbeck, P. & Pezzulo, G. Active inference: a process theory. Neural Comput. 29, 1–49 (2017).
95. Boly, M. et al. Preserved feedforward but impaired top- down processes in the vegetative state. Science 332, 858–862 (2011).
This neuroimaging study uses dynamic causal modelling to show that loss of consciousness in the vegetative state is associated with impaired top- down connectivity from frontal to temporal cortices.
96. Parr, T. & Friston, K. J. Working memory, attention, and salience in active inference. Sci. Rep. 7, 14678 (2017).
97. Chalmers, A. What is This Thing Called Science? (Queensland Univ. Press, 2013).
98. Godfrey- Smith, P. G. Theory and Reality: An Introduction to the Philosophy of Science 2nd edn (Univ. Chicago Press, 2021).
99. Lipton, P. Inference to the Best Explanation (Routledge, 2004).
100. Lau, H. & Passingham, R. E. Relative blindsight in normal observers and the neural correlate of visual consciousness. Proc. Natl Acad. Sci. USA 103, 18763–18768 (2006).
This empirical study compares conscious and unconscious visual perception in humans, controlling for performance, and reveals differences in prefrontal activation.
101. van Vugt, B. et al. The threshold for conscious report: signal loss and response bias in visual and frontal cortex. Science 360, 537–542 (2018).
This empirical study tracks the time course of neural signals in primate frontal cortex, showing that perceived stimuli elicit sustained activity, when compared with non- perceived stimuli.
102. Gaillard, R. et al. Converging intracranial markers of conscious access. PLoS Biol. 7, e61 (2009).
103. Panagiotaropoulos, T. I., Deco, G., Kapoor, V. & Logothetis, N. K. Neuronal discharges and gamma oscillations explicitly reflect visual consciousness in the
lateral prefrontal cortex. Neuron 74, 924–935 (2012).
104. Kapoor, V. et al. Decoding internally generated transitions of conscious contents in the prefrontal cortex without subjective reports. Nat. Comm. 13, 1535 (2022).
105. Bellet, J. et al. Decoding rapidly presented visual stimuli from prefrontal ensembles without report nor post- perceptual processing. Neurosci. Conscious.
2022, niac005 (2022).
106. Levinson, M., Podvalny, E., Baete, S. H. & He, B. J. Cortical and subcortical signatures of conscious object recognition. Nat. Commun. 12, 2930 (2021).
107. Boly, M. et al. Are the neural correlates of consciousness in the front or in the back of the cerebral cortex? Clinical and neuroimaging evidence. J. Neurosci. 37, 9603–9613 (2017).
108. Raccah, O., Block, N. & Fox, K. C. R. Does the prefrontal cortex play an essential role in consciousness? Insights from intracranial electrical stimulation of the human brain. J. Neurosci. 41, 2076–2087 (2021).
109. Odegaard, B., Knight, R. T. & Lau, H. Should a few null findings falsify prefrontal theories of conscious perception? J. Neurosci. 37, 9593–9602 (2017).
110. Brascamp, J., Blake, R. & Knapen, T. Negligible fronto- parietal BOLD activity accompanying unreportable switches in bistable perception.
Nat. Neurosci. 18, 1672–1678 (2015).
This empirical ‘no- report’ study shows that fronto- parietal activity does not track switches in perceptual dominance when subjective reports are not required.
111. Sergent, C. et al. Bifurcation in brain dynamics reveals a signature of conscious processing independent of report. Nat. Commun. 12, 1149 (2021).
112. Siclari, F. et al. The neural correlates of dreaming. Nat. Neurosci. 20, 872–878 (2017).
113. Wong, W. et al. The Dream Catcher experiment: blinded analyses failed to detect markers of dreaming consciousness in EEG spectral power. Neurosci. Conscious. 2020, niaa006 (2020).
114. Block, N. Consciousness, accessibility, and the mesh between psychology and neuroscience. Behav. Brain Sci. 30, 481–548 (2007).
This work argues that research in psychology and neuroscience shows that there is a real and not merely conceptual distinction between phenomenal consciousness (that is, experience) and cognitive access to phenomenal consciousness.
115. Musgrave, A. in Relativism and Realism in Science (ed Nola, R.) 229–252 (Kluwer, 1988).
116. Song, C., Haun, A. M. & Tononi, G. Plasticity in the structure of visual space. eNeuro https://doi.org/ 10.1523/ENEURO.0080-17.2017 (2017).
117. Marshel, J. H. et al. Cortical layer- specific critical dynamics triggering perception. Science https:// doi.org/10.1126/science.aaw5202 (2019).
118. Dembski, C., Koch, C. & Pitts, M. Perceptual awareness negativity: a physiological correlate of sensory consciousness. Trends Cogn. Sci. 25, 660–670 (2021).
119. Sanchez, G., Hartmann, T., Fusca, M., Demarchi, G. & Weisz, N. Decoding across sensory modalities reveals common supramodal signatures of conscious perception. Proc. Natl Acad. Sci. USA 117, 7437–7446 (2020).
120. Sergent, C. The offline stream of conscious representations. Philos. Trans. R. Soc. Lond. B Biol. Sci. https://doi.org/10.1098/rstb.2017.0349 (2018).
121. Michel, M. & Doerig, A. A new empirical challenge for local theories of consciousness. Mind Lang. https://doi.org/10.1111/mila.12319 (2021).
122. Sergent, C. et al. Cueing attention after the stimulus is gone can retrospectively trigger conscious perception. Curr. Biol. 23, 150–155 (2013).
This empirical study reveals that conscious perception of a stimulus can be influenced by events happening (hundreds of milliseconds) after the stimulus appeared (‘retro- perception’).
123. Roseboom, W. et al. Activity in perceptual classification networks as a basis for human subjective time perception. Nat. Commun. 10, 267 (2019).
124. Kent, L. & Wittmann, M. Special Issue: Consciousness science and its theories. Time consciousness: the missing link in theories of consciousness. Neurosci. Conscious. 2021, niab011 (2021).
125. Husserl, E. Ideas: A General Introduction to Pure Phenomenology (Collier Books, 1963).
126. Yaron, I., Melloni, L., Pitts, M. & Mudrik, L. The ConTraSt database for analyzing and comparing empirical studies of consciousness theories. Nat. Hum. Behav. https://doi.org/10.1038/s41562-021-01284-5 (2022).
This work presents an online resource of empirical studies of consciousness, organized with respect to different ToCs.
127. Joglekar, M. R., Mejias, J. F., Yang, G. R. & Wang, X. J. Inter- areal balanced amplification enhances signal propagation in a large- scale circuit model of the
primate cortex. Neuron 98, 222–234.e8 (2018).
128. VanRullen, R. & Kanai, R. Deep learning and the global workspace theory. Trends Neurosci. 44, 692–704 (2021).
129. Shea, N. & Frith, C. D. The global workspace needs metacognition. Trends Cogn. Sci. 23, 560–571 (2019).
130. Suzuki, K., Roseboom, W., Schwartzman, D. J. & Seth, A. K. A deep- dream virtual reality platform for studying altered perceptual phenomenology. Sci. Rep. 7, 15982 (2017).
131. Vilas, M. G., Auksztulewicz, R. & Melloni, L. Active inference as a computational framework for consciousness. Rev. Philos. Psychol. https://doi.org/10.1007/s13164-021-00579-w (2021).
132. Browning, H. & Veit, W. The measurement problem in consciousness. Philos. Top. 48, 85–108 (2020).
133. Seth, A. K., Dienes, Z., Cleeremans, A., Overgaard, M. & Pessoa, L. Measuring consciousness: relating behavioural and neurophysiological approaches. Trends Cogn. Sci. 12, 314–321 (2008).
134. Michel, M. Calibration in consciousness science. Erkenntnis https://doi.org/10.1007/s10670-021-00383-z (2021).
135. Birch, J., Schnell, A. K. & Clayton, N. S. Dimensions of animal consciousness. Trends Cogn. Sci. 24, 789–801 (2020).
136. Bayne, T., Seth, A. K. & Massimini, M. Are there islands of awareness? Trends Neurosci. 43, 6–16 (2020).
This work presents an examination of the possibility of consciousness in isolated neural
systems such as brain organoids, disconnected cortical hemispheres and ex cranio brains.
137. Dehaene, S., Lau, H. & Kouider, S. What is consciousness, and could machines have it? Science 358, 486–492 (2017).
138. Hu, H., Cusack, R. & Naci, L. Typical and disrupted brain circuitry for conscious awareness in full- term and pre- term infants. (2021).
139. Owen, A. M. & Coleman, M. R. Detecting awareness in the vegetative state. Ann. N Y Acad. Sci. 9, 130–138 (2008).
140. Cleeremans, A. The radical plasticity thesis: how the brain learns to be conscious. Front. Psychol. 2, 86 (2011).
141. Jackendoff, R. Consciousness and the Computational Mind (MIT Press, 1987).
142. Prinz, J. The Conscious Brain: How Attention Engenders Experience (Oxford Univ. Press, 2012).
143. Chang, A. Y. C., Biehl, M., Yu, Y. & Kanai, R. Information closure theory of consciousness. Front. Psychol. 11, 1504 (2020).
144. Tononi, G. & Edelman, G. M. Consciousness and complexity. Science 282, 1846–1851 (1998).
This work presents an early proposal of how measures of neural complexity might relate to phenomenological properties of (all) conscious experiences.
145. Edelman, G. M. Neural Darwinism: The Theory of Neuronal Group Selection (Basic Books 1987).
146. Edelman, G. M. The Remembered Present (Basic Books, 1989).
147. Damasio, A. The Feeling of What Happens: Body and Emotion in the Making of Consciousness (Harvest Books, 2000).
148. Graziano, M. S. A. The attention schema theory: a foundation for engineering artificial consciousness. Front. Robot. AI 4, 60 (2017).
149. Dennett, D. C. Consciousness Explained (Little, Brown, 1991).
150. Ginsburg, S. & Jablonka, E. The Evolution of the Sensitive Soul: Learning and the Origins of Consciousness (MIT Press, 2019).
151. Aru, J., Suzuki, M. & Larkum, M. E. Cellular mechanisms of conscious processing. Trends Cogn. Sci. 24, 814–825 (2020).
152. McFadden, J. Integrating information in the brain’s EM field: the cemi field theory of consciousness. Neurosci. Conscious. 2020, niaa016 (2020).
153. Fleming, S. M., Ryu, J., Golfinos, J. G. & Blackmon, K. E. Domain- specific impairment in metacognitive accuracy following anterior prefrontal lesions. Brain 137, 2811–2822 (2014).
154. Fox, K. C. R. et al. Intrinsic network architecture predicts the effects elicited by intracranial electrical stimulation of the human brain. Nat. Hum. Behav. 4, 1039–1052 (2020).
155. Dehaene, S. & Naccache, L. Towards a cognitive neuroscience of consciousness: basic evidence and a workspace framework. Cognition 79, 1–37 (2001).
156. Sergent, C., Baillet, S. & Dehaene, S. Timing of the brain events underlying access to consciousness during the attentional blink. Nat. Neurosci. 8, 1391–1400 (2005).
157. Mediano, P. A. M., Seth, A. K. & Barrett, A. B. Measuring integrated information: comparison of candidate measures in theory and simulation. Entropy 21, 17 (2019).
158. Casali, A. G. et al. A theoretically based index of consciousness independent of sensory processing and behavior. Sci. Transl. Med. 5, 198ra105 (2013).
This empirical study shows that a measure of the complexity of the cortical response to transcranial magnetic stimulation distinguishes between a range of global conscious states, including disorders of consciousness.
159. Luppi, A. I. et al. Consciousness- specific dynamic interactions of brain integration and functional diversity. Nat. Commun. 10, 4616 (2019).
160. Hardstone, R. et al. Long- term priors influence visual perception through recruitment of long- range feedback. Nat. Commun. 12, 6288 (2021).
161. de Lange, F. P., Heilbron, M. & Kok, P. How do expectations shape perception? Trends Cogn. Sci. 22, 764–779 (2018).
162. Melloni, L., Schwiedrzik, C. M., Muller, N., Rodriguez, E. & Singer, W. Expectations change the signatures and timing of electrophysiological correlates of perceptual awareness. J. Neurosci. 31, 1386–1396 (2011).
This empirical study uses a perceptual hysteresis paradigm to show that expectations enhance and accelerate conscious perception.
163. Pinto, Y., van Gaal, S., de Lange, F. P., Lamme, V. A. & Seth, A. K. Expectations accelerate entry of visual stimuli into awareness. J. Vis. 15, 13 (2015).
164. Chalmers, D. J. Facing up to the problem of consciousness. J. Conscious. Stud. 23, 200–219 (1995).
This work presents the classic statement of the philosophical distinction between the ‘hard’ and ‘easy’ problems of consciousness.
165. Levine, J. Materialism and qualia: the explanatory gap. Pac. Philos. Q. 64, 354–361 (1983).
166. Seth, A. K. The Real Problem (Aeon, 2016
167. Balog, K. in The Oxford Handbook of Philosophy of Mind (eds Beckermann, A., McLaughlin, B. P., & Walter S.) 292–312 (Oxford Univ. Press, 2009).
168. Perry, J. Knowledge, Possibility, and Consciousness (MIT Press, 2001).
169. Varela, F. J., Thompson, E. & Rosch, E. The Embodied Mind: Cognitive Science and Human Experience (MIT Press, 1993).
170. Carvalho, G. B. & Damasio, A. Interoception and the origin of feelings: a new synthesis. Bioessays 43, e2000261 (2021).
171. Solms, M. The Hidden Spring: A Journey to the Source of Consciousness (Profile Books, 2021).
172. Merker, B. Consciousness without a cerebral cortex: a challenge for neuroscience and medicine. Behav. Brain Sci. 30, 63–81; discussion 81–134 (2007).
173. Parvizi, J. & Damasio, A. Consciousness and the brainstem. Cognition 79, 135–160 (2001).
174. Naber, M., Frassle, S. & Einhauser, W. Perceptual rivalry: reflexes reveal the gradual nature of visual awareness. PLoS ONE 6, e20910 (2011).
175. Casarotto, S. et al. Stratification of unresponsive patients by an independently validated index of brain complexity. Ann. Neurol. 80, 718–729 (2016).
176. Shea, N. & Bayne, T. The vegetative state and the science of consciousness. Br. J. Philos. Sci. 61, 459–484 (2010).
177. Birch, J. The search for invertebrate consciousness. Noûs 56, 133–153 (2020).
178. Phillips, I. The methodological puzzle of phenomenal consciousness. Philos. Trans. R. Soc. Lond. B Biol. Sci. https://doi.org/10.1098/rstb.2017.0347 (2018).
(参考文献可上下滑动查看)
未来智能实验室的主要工作包括:建立AI智能系统智商评测体系,开展世界人工智能智商评测;开展互联网(城市)大脑研究计划,构建互联网(城市)大脑技术和企业图谱,为提升企业,行业与城市的智能水平服务。每日推荐范围未来科技发展趋势的学习型文章。目前线上平台已收藏上千篇精华前沿科技文章和报告。
如果您对实验室的研究感兴趣,欢迎加入未来智能实验室线上平台。扫描以下二维码或点击本文左下角“阅读原文”



