
- 1matlab ~的用法_matlab d~
- 2Java selenium使用ChromeDriver截图 解决get超时后续任务报错问题_chromedriver setscripttimeout
- 3android上改变listView的选中颜色_android listview选中变色
- 4基本select使用 全解
- 5安装rpm包冲突_conflicts with file from package filesystem-3.2-25
- 6SLAM数据集【百度网盘】_tum-rgbd数据集百度网盘下载
- 7[libco] 协程库学习,测试连接 mysql_作为服务底层
- 8二维有限元的MATLAB仿真_fpga教程
- 9为什么Kubernetes已经成为程序员必备技能_java开发需要学k8s吗
- 10Iterator(迭代器)的用法及其背后机制的探究_this.iterator()
文献阅读-10X单细胞揭示肿瘤浸润性T细胞的泛癌单细胞图谱_单细胞测序odds ratio
赞
踩
Pan-cancer single cell landscape of tumor-infiltrating T cells
INTRODUCTION
针对肿瘤特异性T细胞的癌症免疫疗法使许多癌症患者受益,但不同的癌症类型的临床疗效差异很大。肿瘤浸润性T细胞通常会进入功能失调状态,即众所周知的Tex,effector T cells,的功能受多种因素的调节,包括调节性的T细胞,T细胞的状态和丰度因不同癌症类型的肿瘤微环境(TME)而异,这可能从根本上影响不同的临床参数,例如对免疫疗法的药物反应。
RATIONALE
为了建立高分辨率的pan-Cancer的T细胞atlas,作者对来自不同病人tumor,癌旁组织以及血液样本进行单细胞测序,并收集其他已经发表的单细胞测序的数据集。在矫正混杂因素和批次效应以后整合不同的数据集。该图谱来自316名病人共21种癌症类型。将具有基因表达谱的单个T细胞的T细胞受体序列组装起来,以表征T细胞的扩增和动态(expansion and dynamics)。不同统计学的方法来研究不同癌症类型的特征和丰度。
RESULTS
在癌症患者中发现了多个潜在pTRT(potentially tumor-reactive T cell)的类型。pTRTs的状态在不同的肿瘤微环境中变化很大。
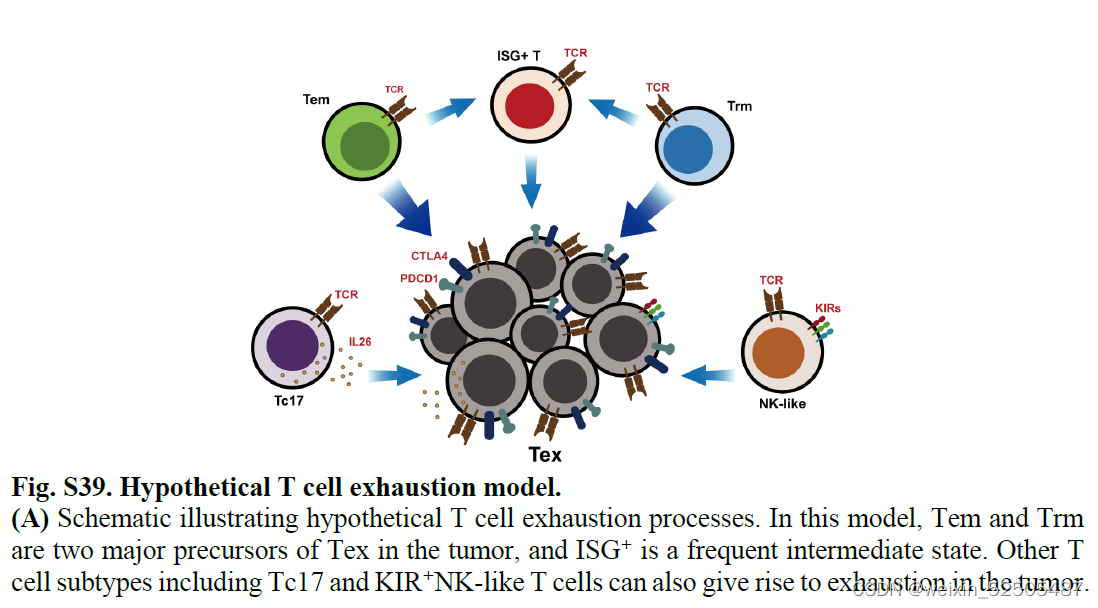
对于CD8+ T细胞来说,主要的pTRTs是exhausted T cells并表现出很高的异质性。作者通过计算推断出exhausted T cells两条主要的发育途径,包括通过effector memory T cells和tissue-resident memeory T cells发育为exhausted T cells。并且证明了这两条耗竭的通路在不同类型的癌症中十分普遍。作者同时注意到了terminal exhausted T cells 和其他的细胞如natural killer (NK)-like T cells, Tc17和Treg细胞之间的状态转化。但是这种转化往往发生在特定的癌症类型中。
对于CD4+ T细胞,如follicular helper T cell (TFH)/T helper(TH1)双功能的T细胞可能是TFH细胞转化而来。显著的pTRTs与TMB相关。作者还发现TME中的TGF-β和IFN可能会影响pTRTs的转录程序。T细胞状态的丰度因癌症的类型而定。根据肿瘤浸润性T细胞的组成,可以将癌症患者的免疫类型分为含量较高的terminal exhausted T cells以及具有高频的tissue-resident memory T cells。患者的免疫表型与临床症状例如患者的生存和免疫检查点阻断剂的反应
Abstract
T细胞在肿瘤免疫治疗中扮演重要的角色,但是缺乏对不同癌症类型浸润性T细胞的异质性和动态性的系统性的比较。作者通过建立316名患者包括21种癌症类型和pan-cancer的单细胞测序,揭示了不同的T细胞组成模式。作者发现在exhaustion T cells 中多条细胞状态转化通路并且细胞类型的转化存在于多种癌症类型中,某些T细胞群显示出于患者特性(如肿瘤突变负荷)的相关性。揭示了肿瘤微环境可能的决定因素(shedding light on the possible determinants of the tumor microenvironment)。仅肿瘤内T细胞的组成就可以将患者分为具有不同临床特性的组,为T细胞的免疫治疗以及靶向T细胞的精准治疗提供新的见解。
INTRODUCTION
TILs是肿瘤浸润免疫微环境中的核心参与者,塑造了基本的临床症状如对免疫治疗的反应。免疫检查点阻断剂已经显示出巨大的临床治疗成功(tremendous),但是治疗效果在不同的癌症类型之间存在巨大的差异,这表明免疫治疗存在潜在的差异。在TME中,effector T cells往往高表达抑制性受体如PD-1,TIM3,TIGIT,LAG3,她们往往是功能失调的标志,被广泛称为exhausting T cells。ICB的功效可能与不同类型癌症之间的TILs的状态有关,尤其是exhausting T cells 的差异。在黑色素瘤患者中,CD8+ 肿瘤浸润性T细胞表现出从前功能障碍细胞状态到功能障碍细胞状态的线性和连续的发展。但是在肺癌的患者体内,有两种耗竭前的状态可以发展为exhausting状态,因此,各个癌症TME之间耗竭动态可能不同。从本质上讲(Intrinsically),T细胞耗竭的发生可能与几种TFs相关,包括TOX,TCF7,以及形成在在功能失调的CD8+ T细胞群的表观遗传的调控因子。除此之外,多种TME相关的因子也可能会导致耗竭的表型,并且在各种癌症类型的TME中可能存在决定T细胞表型和丰度的不同调节过程。在不同的癌症类型中观察到了可以区分T细胞特征。例如,在肝细胞癌和结直肠癌中exhausting T cells的占比比肺癌中高。多发性骨髓瘤等没有显著的exhausted T cells的亚群。但是,直接的比较研究仅限于三四种孤立的癌症类型。
通过单细胞测序作者构建了跨21种不同癌症类型的综合肿瘤浸润T细胞纲要(compendium),通过寻找TILs的共同点和差异,我们揭示了T细胞状态,动力学和调节的pan-cancer特性。
RESULTS
construction of a pan-cancer single-cell transcrptome atlas of T cells
作者表征了21种癌症类型的T细胞转录谱。Figure1A Schematics of pan-cancer single-cell transcriptome and TCR profiling of T cells
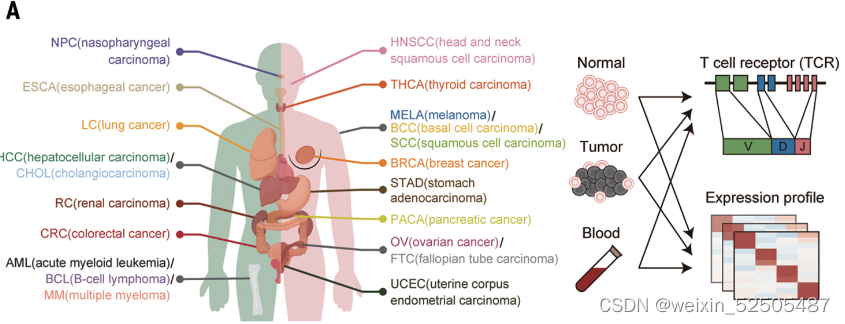
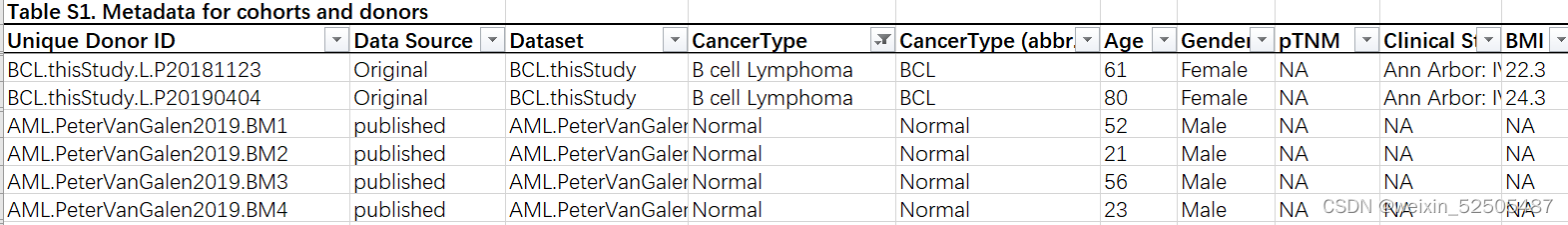
经过严格的质量控制以后,包含来自316个供体的397810个T细胞的数据-来自肿瘤,临近的正常组织以及外周血。其中46.6的细胞是本研究新测序的细胞,其他的数据则来自于先前测序的数据集。tableS1
作者在微型集群和Harmony矫正批次效应的基础上结合先前研究的多项数据 FigureS1 FigureS2 FigureS3
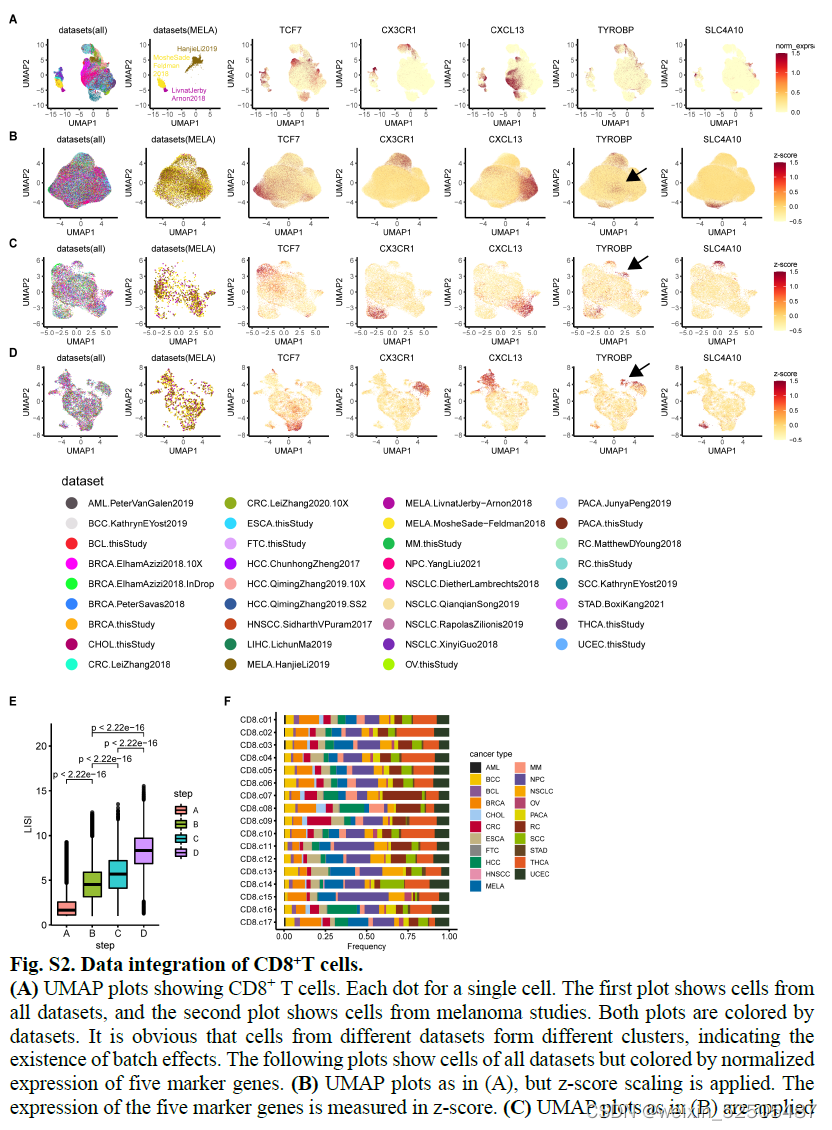

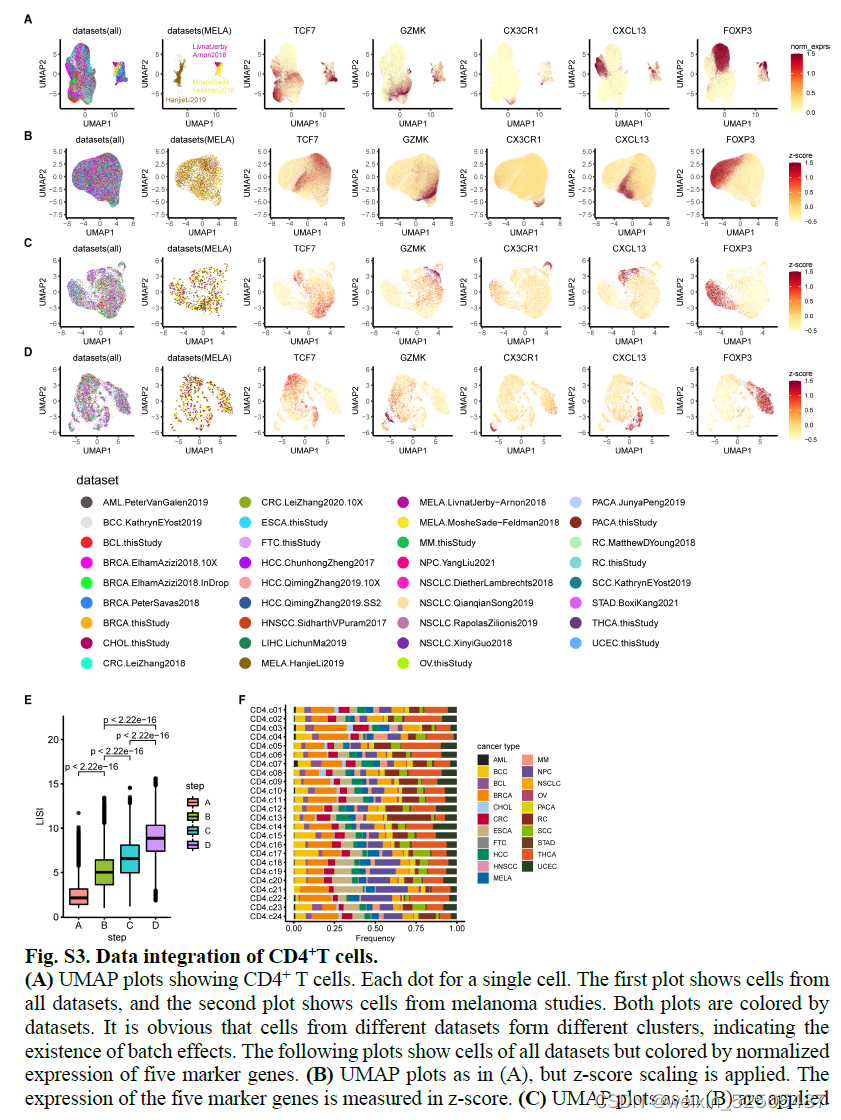

在图像以及LISI结果上表明多项数据混合以后效果很好FigureS2E,FigureS3E
通过10x VDJ and Smart-Seq2测序得到的单个T细胞的TCR结果被assembled。
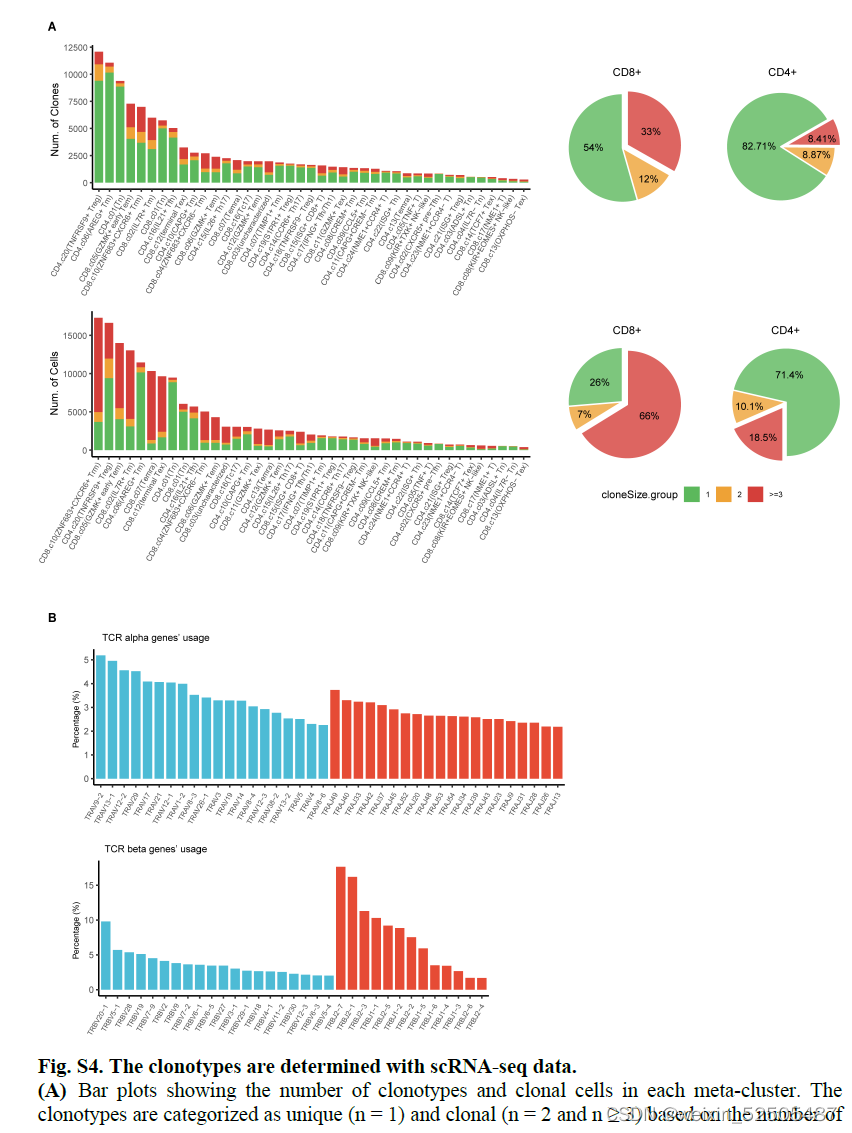
168,901 cells from 92,533 clonotypes spanning 87 donors from 15 cancer types来自15种癌症类型的87个供体的92,533个克隆型共168,901个细胞具有至少一对生产性TCR α chain and β chain,其中53.9%是克隆细胞(在至少两个细胞中发现相同的TCR对),对应14,631扩展克隆亚型。FigureS4

总共确认了17种CD8+ T细胞和24种CD4+T细胞metacluster(Figure1B,C),这些细胞亚型至少在80%的癌症类型中共享(FigureS2F,FigureS3F)。
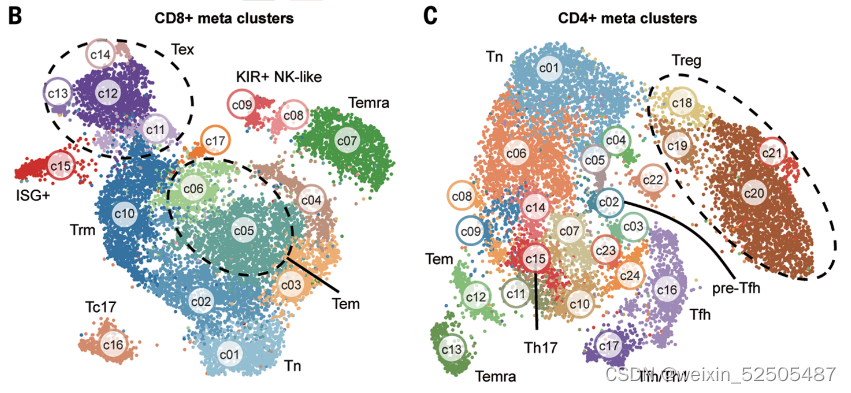
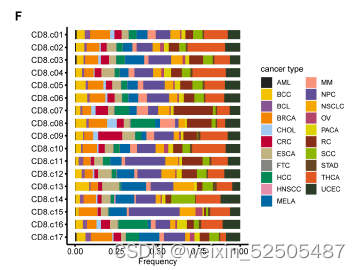
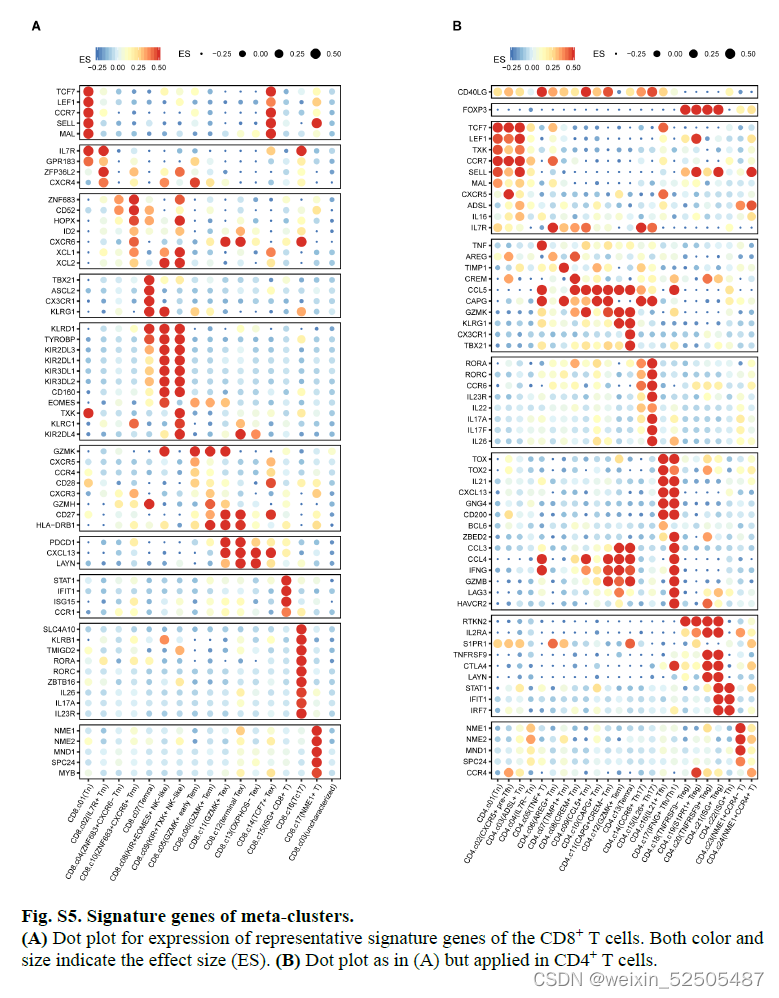
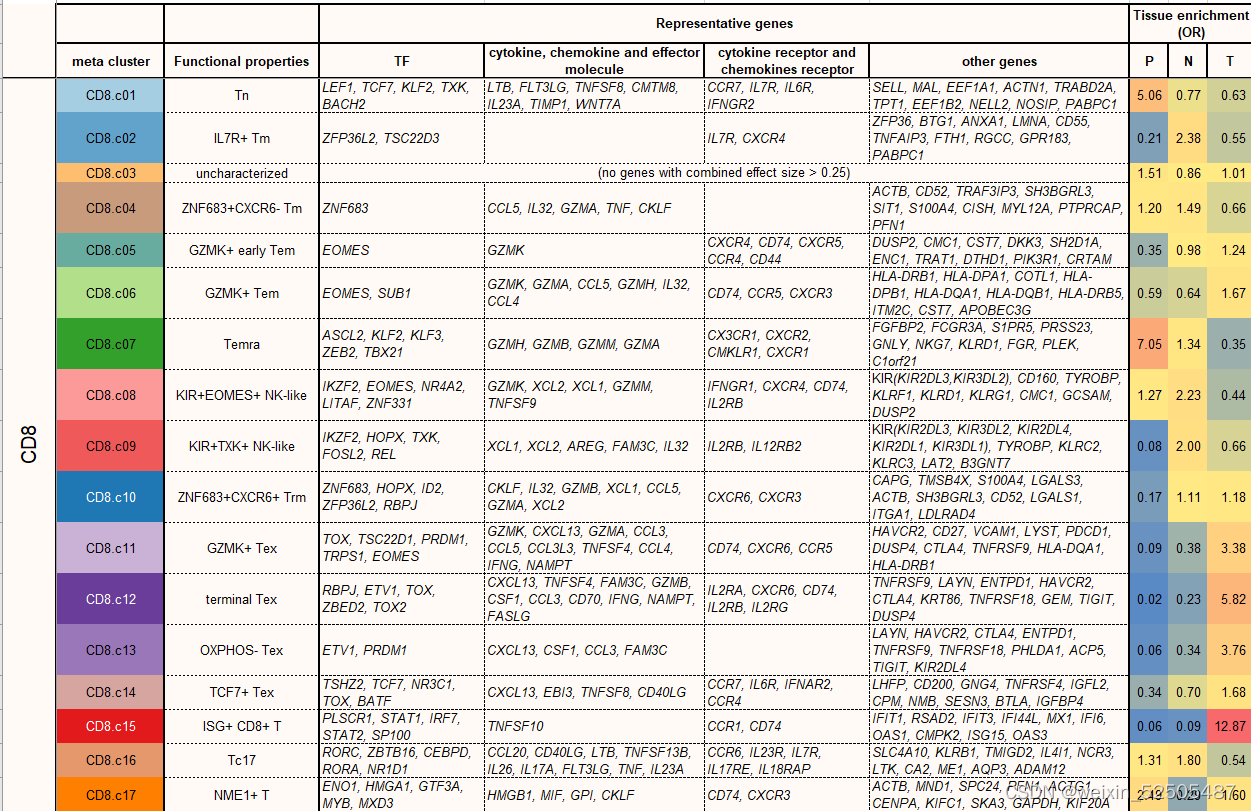
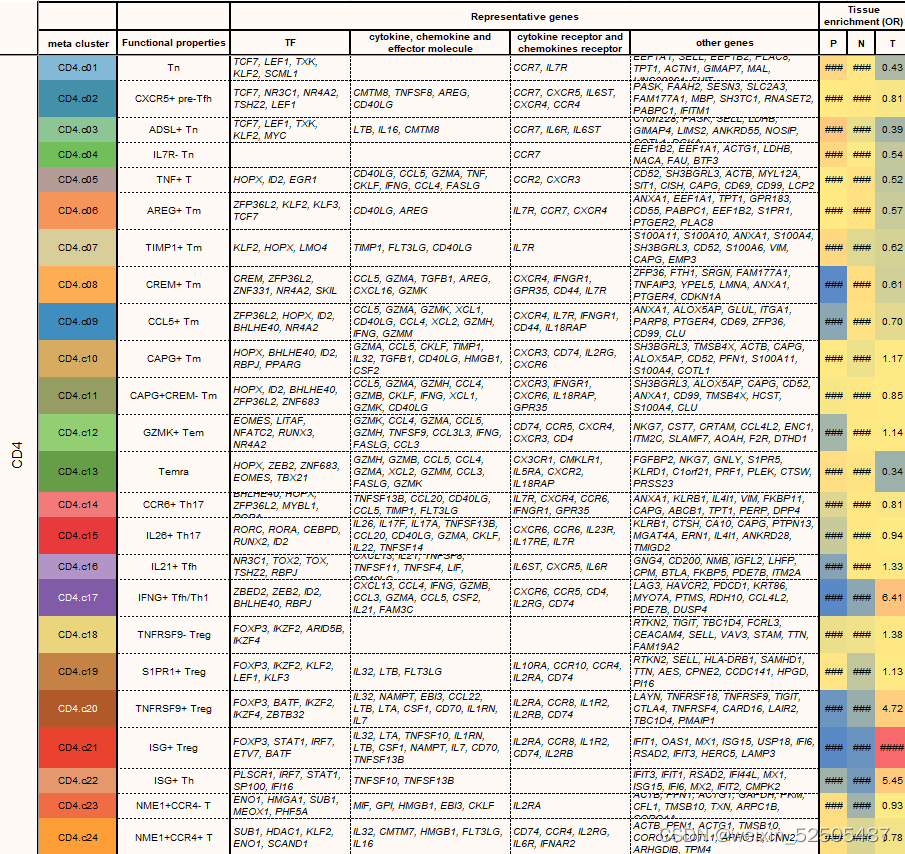
这些表达特征揭示了先前描述的T细胞亚型和新group的出现,包括GZMK+ effector memory T cells(Tem cells)terminally differentiated effector memory or effector cells(Terma)以及interferon-stimulated genes (ISG)-positive T cells in both CD4+ and CD8+ compartments(ISG+ T cells);killer cell immuoglobulin-like receptor(KIR)-postive natural killer(NK)-like T cells(KIR+ NK-like T cells);ZNF683+ CXCR6+ tissue-resident memory T cells (Trm cells)以及四种exhausted CD8+ T cells(Tex cells);三种follicular helper T cell(TFH cell)-related populations(CXCR5+ pre-TFH,classica IL21+ TFH, IFNG+ TFH/T helper (TH1)双功能T细胞)和四种调节性CD4+ T细胞(Treg cell)FigureS5 TableS2
对于CD8+ T细胞c16 metacluster进一半细胞具有半不变的TCR α 链黏膜相关不变T细胞(MAIT)以及有或没有TCR α链细胞均高表达与17型CD8+ T细胞(Tc17细胞)相关的基因,表明这种metacluster包含MAIT或non-MAIT Tc17细胞(一下简称Tc17)。
未经治疗的患者不同来源的T细胞组成表现出显著的差异,因为用Shannon equitability index测量正常组织和肿瘤组织的多样性显著高于血液样本,对于CD8+ T细胞,肿瘤组织中的多样性高于正常组织的多样性。Figure1D and E Figure S7A、B
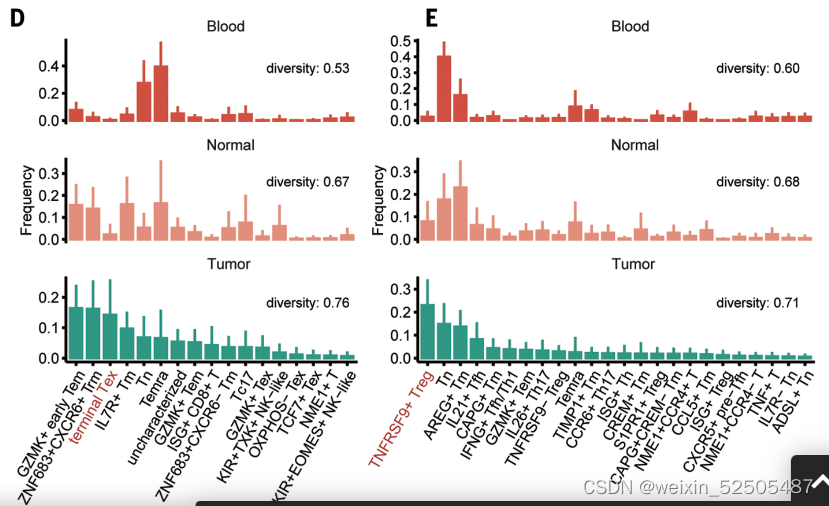
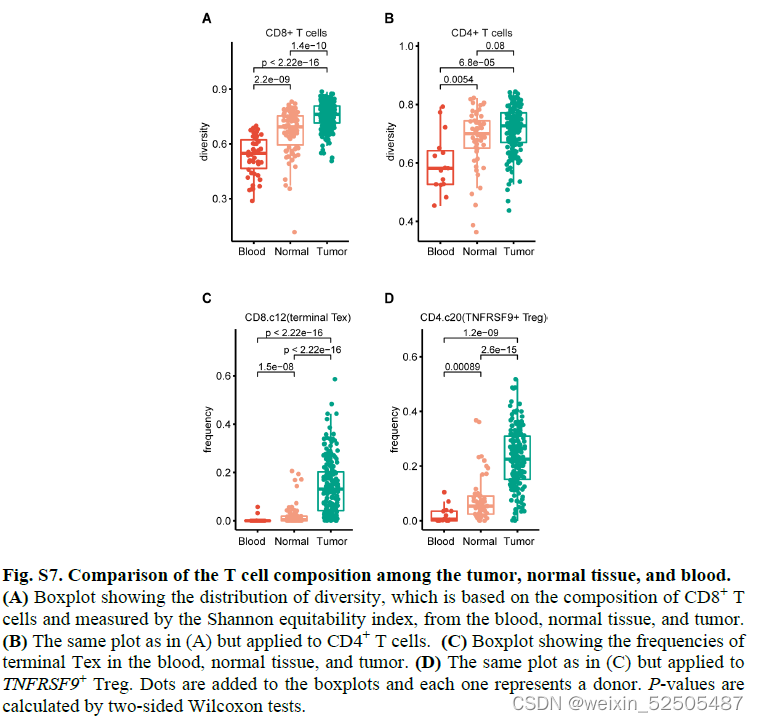
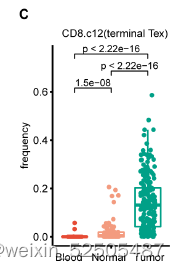
CD8+ T细胞组成主要是以耗竭细胞为特征Tex(Figure 1D, FigS7C)
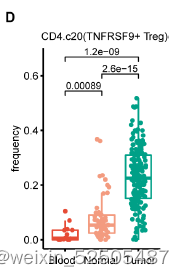
而在CD4+ T细胞中主要是以TNFRSF9+ Treg细胞在肿瘤组织中占比最多,而在血液和正常组织的样本中占比较少。Figure 1E FigureS7D
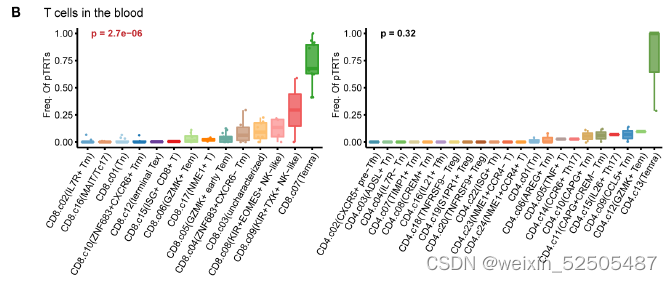
为了鉴别对肿瘤反应性的T细胞以及bystander T cells,作者对组织分布,转录表型,增殖和克隆扩增的特性进行联合分析。富含肿瘤的T细胞的增殖和克隆的扩增特性已经被证明是肿瘤反应性的证据。从odds ratio(OR)分析结果来看,naive T cells和Temra cell(包括CD8+ CD4+ T细胞)在血液中表现为强烈的分布偏好;而TNFRSF9+ Treg细胞,TFH/TH1细胞,CD8+ ISG+ T cells以及四种CD8+ Tex细胞都呈现肿瘤组织富集的特性。Figure1F

在STARTRAC分析的基础上(single T cells analysis by RNA-seq and TCR tracking),例如tumor-enriched metacluster表现出扩张和持续性的增殖,暗示其克隆扩张是对肿瘤抗原的反应。Figure1G
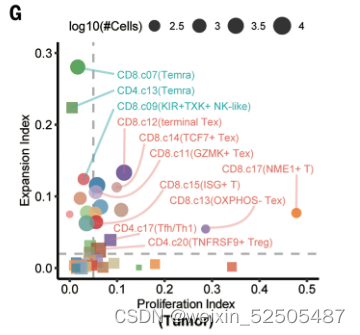
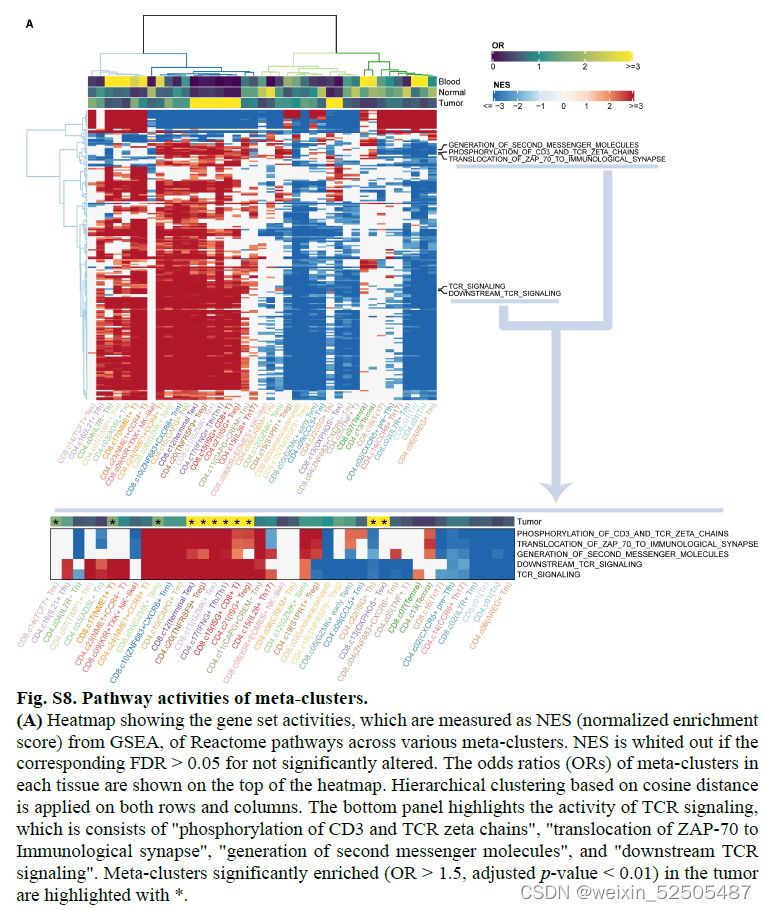
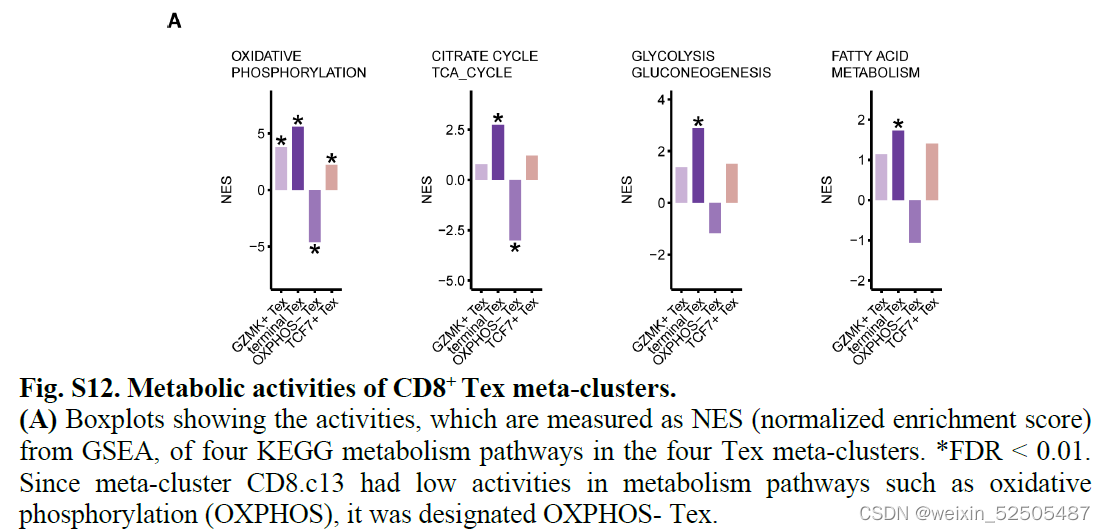
除此之外,大部分具有高扩张和增殖指数的tumor-enriched metaclusters(除了TCF7+ Tex cells 和OXPHOS- Tex cells)趋向于在TCR信号通路上表现为高活性,证实了它们高抗原反应性。FigureS8
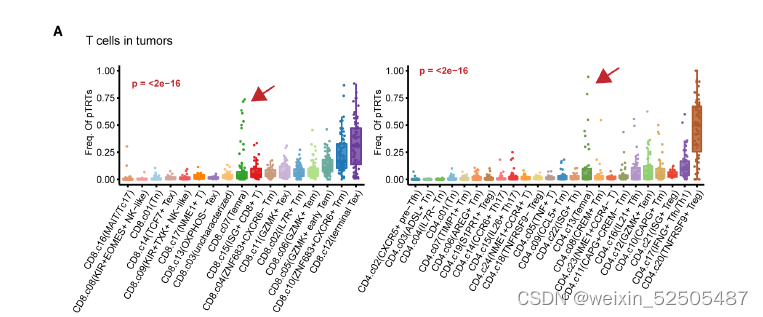
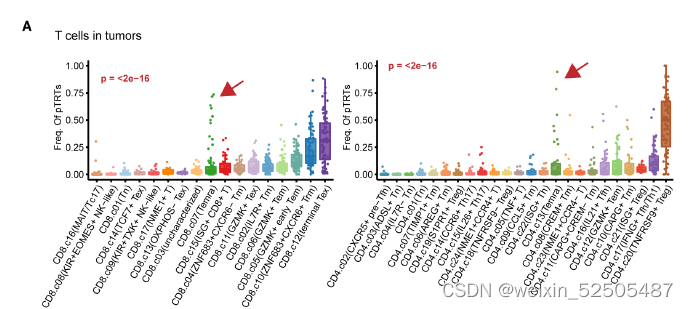
在个体肿瘤水平,作者确定了具有高活性TCR信号通路或高增殖活性的特定扩增的TILs共享克隆型的细胞组,这些细胞被统称为potential tumor-reactive T cells(pTRTs)。对于肿瘤中的pTRTs,对于CD8+ T 和CD4+ T来说最常观察到的细胞状态是terminal Tex cells以及TNFRSF9+ Treg细胞 FigureS9A
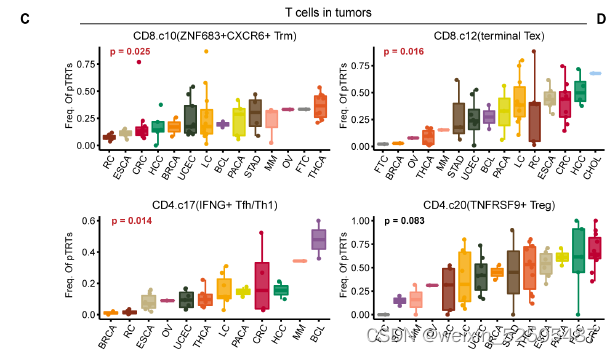
尽管它们的发生率因癌症类型而定,FigureS9C
同时,CD4+和CD8+ compartments 中的Temra cells在所有的组织中表现为显著的扩增(p < 0.01,permutation test),但是增值率较低(<2%)。FigureS1G and Fig.S10
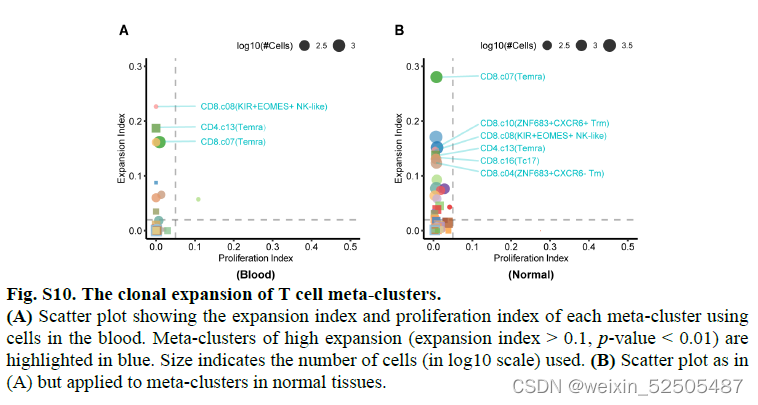
量化组织迁移程度的STARTRAC迁移指数显示,在大多数测试的癌症类型中,CD4+ T 和CD8+ Temra细胞在血液、正常组织、肿瘤组织之间迁移率最高。Figure11A,B
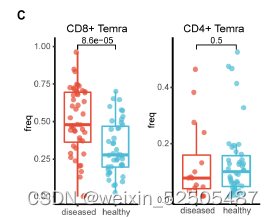
以上结果均表明这些T细胞在肿瘤外被激活和扩增并且在血液中循环。与健康的捐赠者相比,癌症的患者血液中含有更多Temra cells (FigS11C)
此外,在某些肿瘤中,大多数的pTRTs是Temra细胞Fig.S9A
此外,血液中最常见的细胞状态也是Temra细胞 Fig.S9B
并且,这种模式适应于多种癌症。
综上所述,出现了potentially tumor-reactive T cells,TFH/TH1细胞,TNFRSF9+ Treg cells,CD8+ ISG+ T cells以及四种Tex细胞群,代表了局部抗肿瘤反应,而CD8+ Temra细胞可能含有肿瘤特定的TCRs。这与全身免疫反应的概念一致。
Common themes of CD8+ Tex cell heterogeneity and dynamics
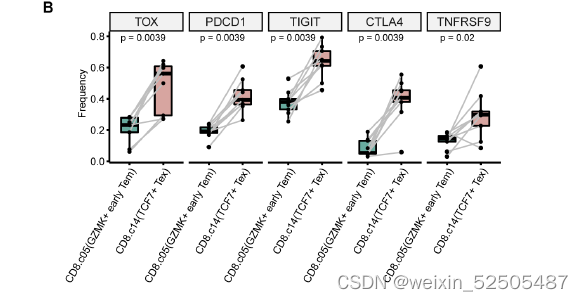
在CD8+ T细胞的亚群中,最主要的pTRTs的metaclusters是四种Tex细胞,所有Tex细胞亚群高表达exhaustion markers,包括TOX,TIGIT,CTLA4以及TNFRSF9。但是它们在基因的表达和通路的活化方面有所不同。Fig.2A and Fig.S12
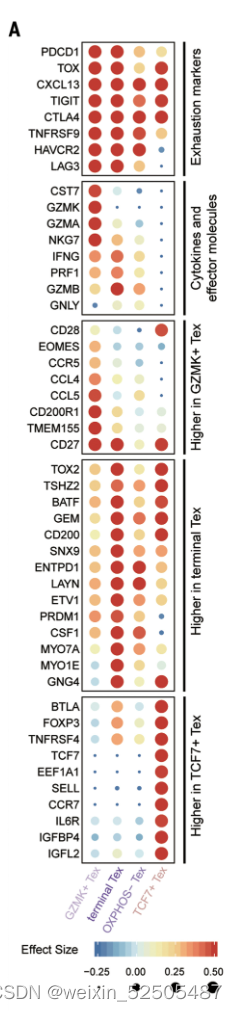
pTRTs的主要亚群为terminal Tex CD8+ T cells,高表达与末端分化相关的基因ENTPD1(外核苷三磷酸二磷酸水解酶 1),还高表达IFNG和GZMB,暗示其内在的在抗肿瘤效应潜力以及某些在T细胞耗竭中具有未知作用的基因,包括MYO7A和MYO7E,相对稀有的Tex群是TCF7+ Tex细胞群,具有低水平的HAVCR2(甲型肝炎病毒细胞受体 2)和LAG3(淋巴细胞活化 3),但是特殊高表达TCF7。TCF7被认为是肿瘤中干细胞样T细胞关键的调节因子。除此之外,TCF7+ Tex高表达CD200,GNG4,IGFBP4和IGFL2以及淋巴结迁移相关基因包括CCR7和SELL。
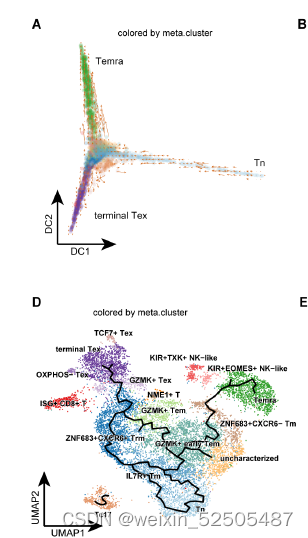
接下来,作者结合基因表达谱信息和TCR数据来剖析T细胞耗竭的轨迹。首先,on a global scale, diffusion map和RNA velocity结果表明CD8+ T细胞从幼稚型T细胞发育为Temra或Tex细胞。Fig.S13A,B,这与先前的报道一致。
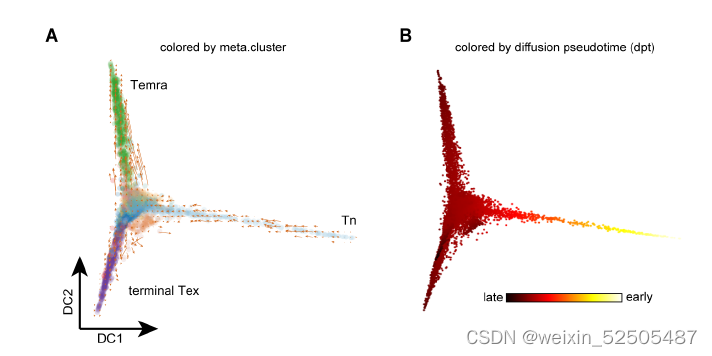

第二,对于Tex分支,结合Monocle3,RNA velocity和graph inference based on uniform manifold approximation and projection (UMAP)以及STARTRAC成对转化索引(pTrans)图推断从Naive T cells到Tex细胞有两条主要的通路,第一条通路 (P1)going through GZMK+ Tem cells [naïve cells to IL7R+ memory T cells (Tm cells) to GZMK+ T cells to terminal Tex cells],第二条通路(P2)going through ZNF683+ Trm cells (naïve cells to IL7R+ Tm cells to ZNF683+CXCR6+ Trm cells to terminal Tex cells)。 Fig.2B, C and C figs. S13, C to E Fig.S14
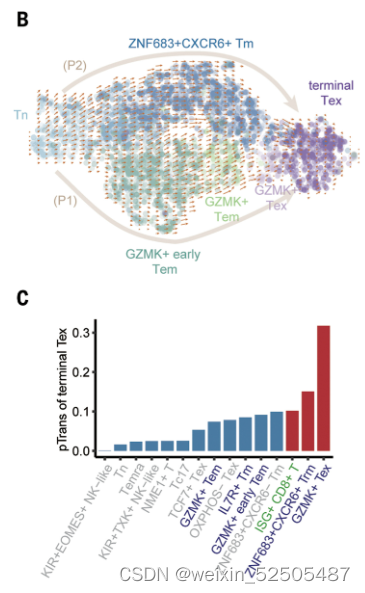
figs. S13, C to E
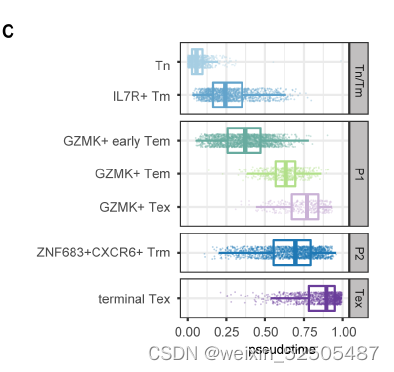
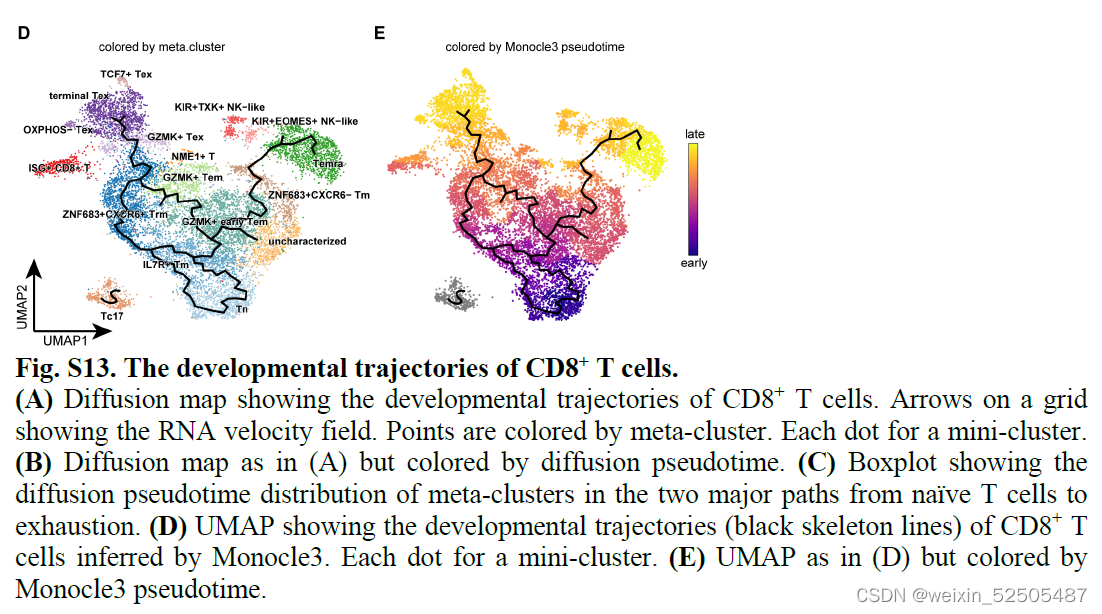
Fig.S14
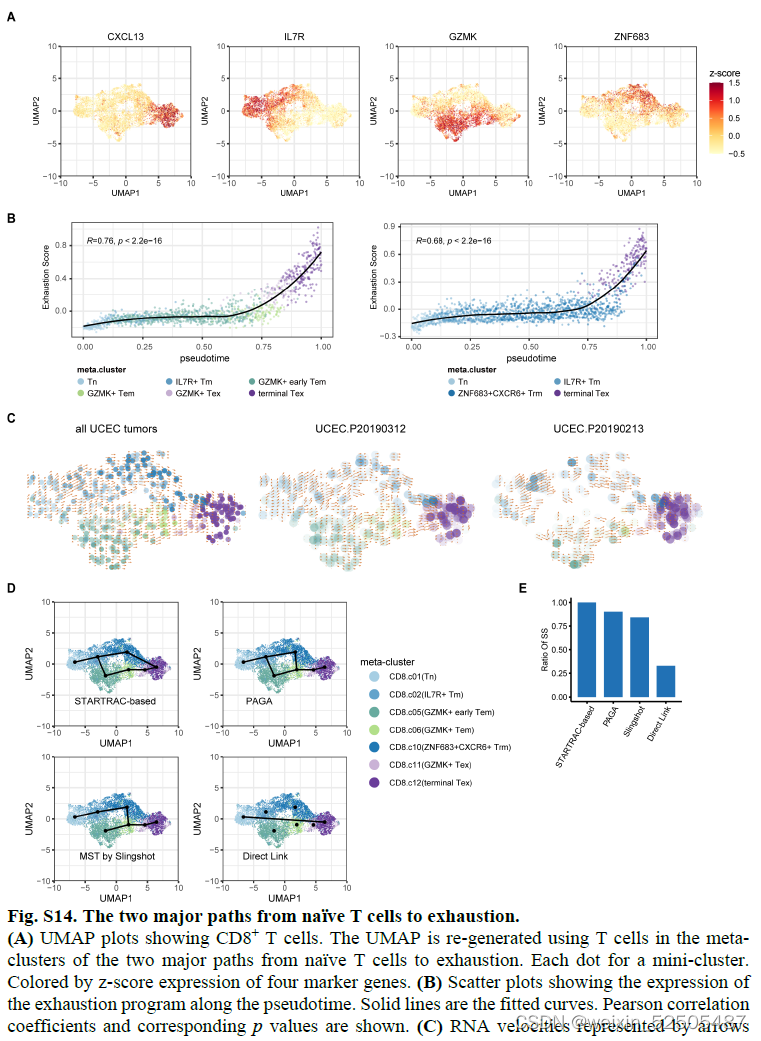

在单个肿瘤中也可以观察到发育到Te细胞群的两条通路 Fig.S15
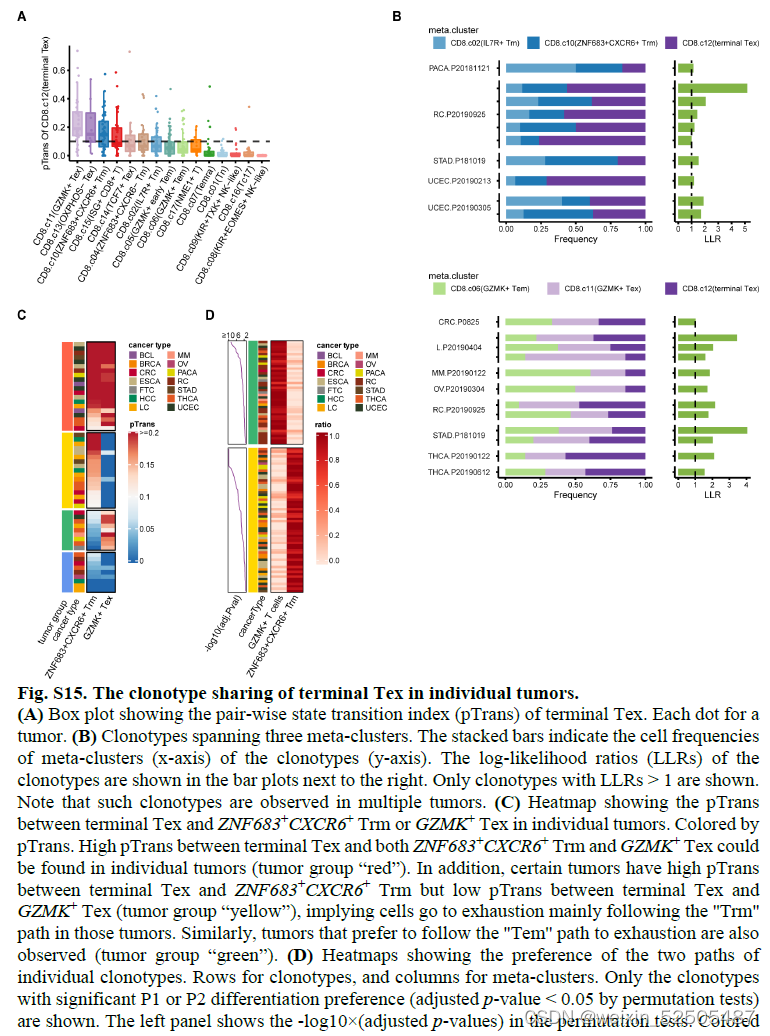

尽管某些肿瘤表现出优先使用P1或P2,但是在其他的肿瘤中,这两条通路的状态转变很高。这意味着Tem和Trm的细胞都参与了抗肿瘤的免疫 Fig.S15C
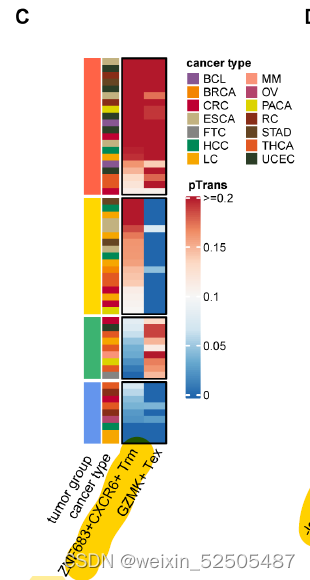
terminal Tex细胞具有和ISG+ T细胞中等的pTrans(the state transition index and the pairwise state transition index (pTrans))。Fig.S16A
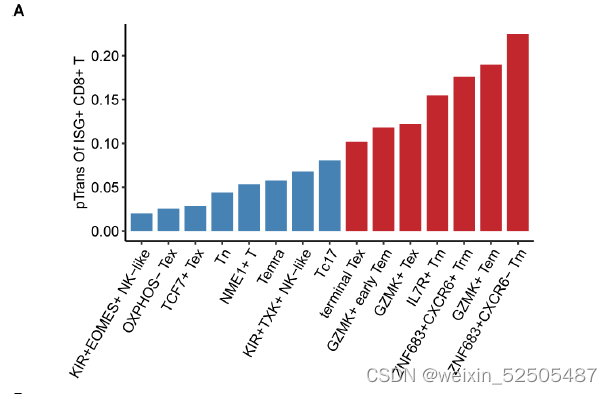
ISG+ T细胞状态并不是一个单独的状态,是一个混合了Tem,Trm和其他细胞。fig. S17
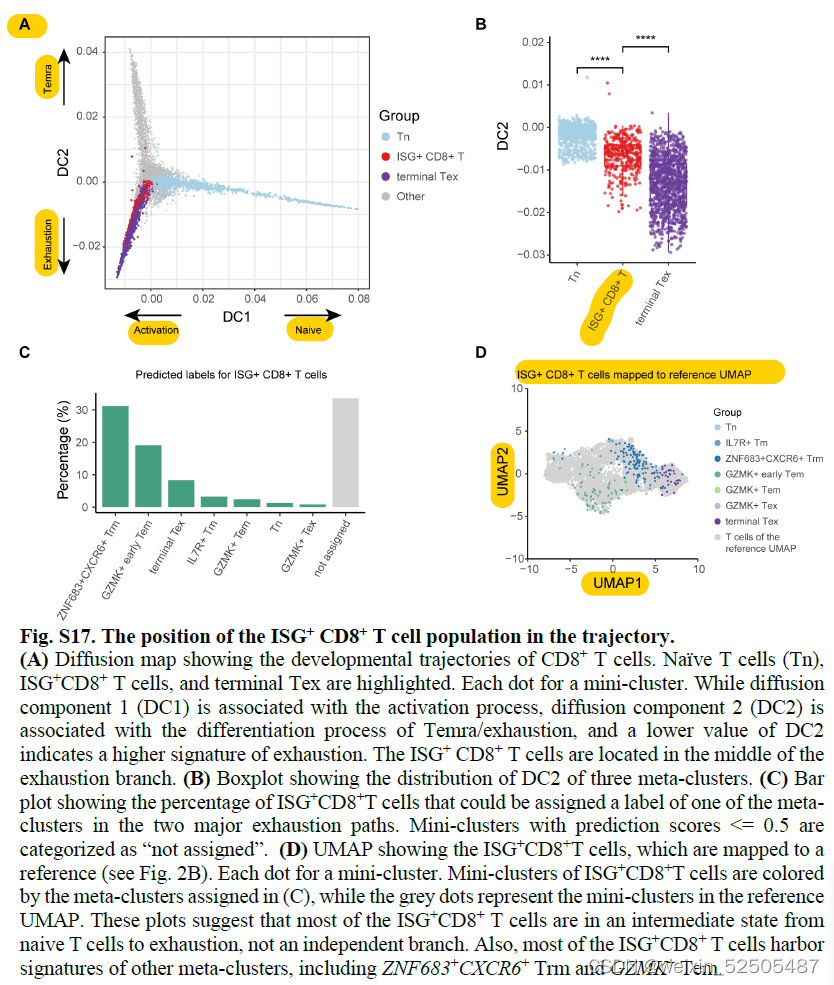
除此之外,包含有多种细胞状态的TCR克隆亚型,包括ZNF683+CXCR6+ Trm cells or GZMK+ Tex cells, ISG+ T cells, and terminal Tex cells可以在肿瘤中被清楚的识别出来。fig. S16, B and C

因为ISG+ 状态代表了可能由TCR触发的interferon-γ (IFN- γ)或直接由干扰素触发的细胞状态。以上的结果表明P1或P2通路中未耗竭的T细胞在进入耗竭状态之前可能会进入到干扰素代表的状态之中。这让人想起在慢性病毒感染的过程中,CD8+ T细胞-内在type Ⅰ干扰素信号使分化偏向于更末端的效应状态或衰竭状态。
假设表达CXCR5或TCF7祖细胞产生terminal Tex细胞,我们发现TCF7+ Tex细胞和GZMK+ Tex cells and even terminal Tex cells有很强的状态转化来连接 fig. S18
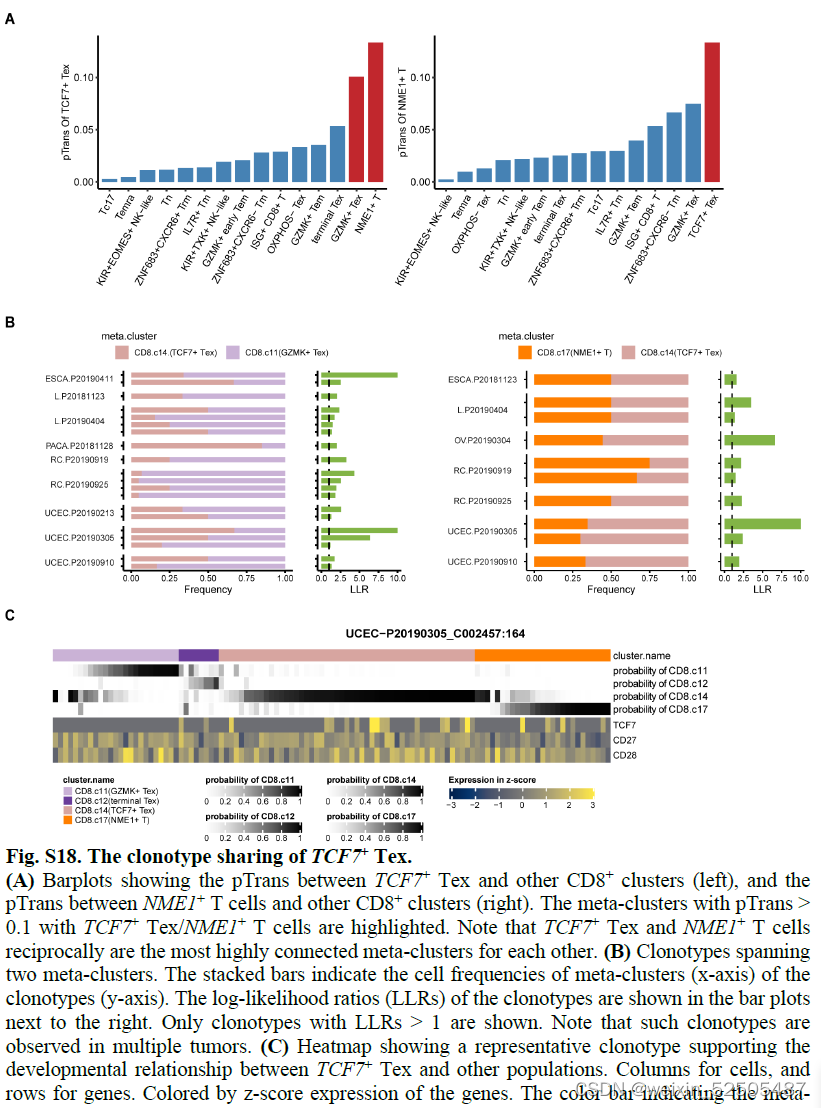
GZMK+ early Tem也表达高水平的TCF7和CXCR5 fig. S19A
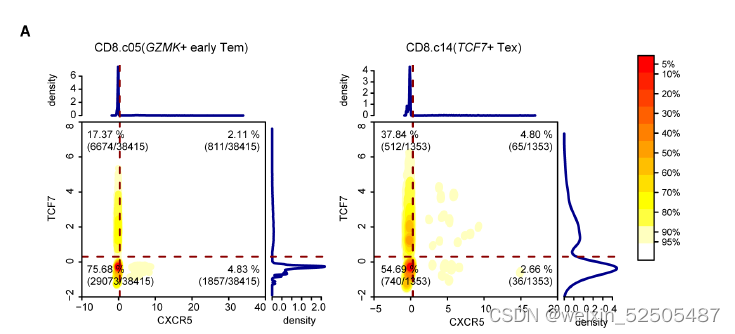
这些来自于GZMK+ early Tem中表达TCF7和CXCR5的细胞群主要位于Temra或Tex分支的附近。fig. S13, A and D 代表具有不同命运的发育潜力的细胞
相反,TCF+ Tex细胞更可能代表一定干性的的Tex细胞,相比于GZMK+ early Tem cells,TCF7+ Tex细胞中TCF7+和CXCR5+的细胞具有更高频率的细胞表达TOX和抑制性受体如PDCD1, TIGIT, and CTLA4。fig. S19B
TCF7+ Tex cells与GZMK+ early Tem没有强烈的状态转变,但是与 NME1+ (NME/NM23 nucleoside diphosphate kinase 1) T cells高度相关 figs. S18
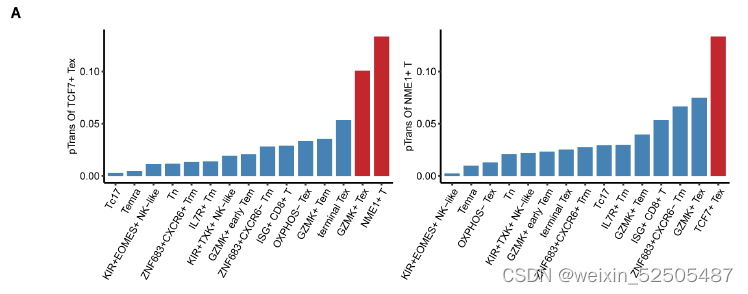
由于这两种细胞群的稀缺性(scarcity),TCF7+ Tex cells是否来自于NME1+ cells还有待于观察。
因此,作者确定了不能单独用CXCR5或TCF7定义的T细胞耗竭途径,总之,作者他们确定了一条更加复杂的耗竭途径。
Distinct paths to T cells exhaustion across different cancer types
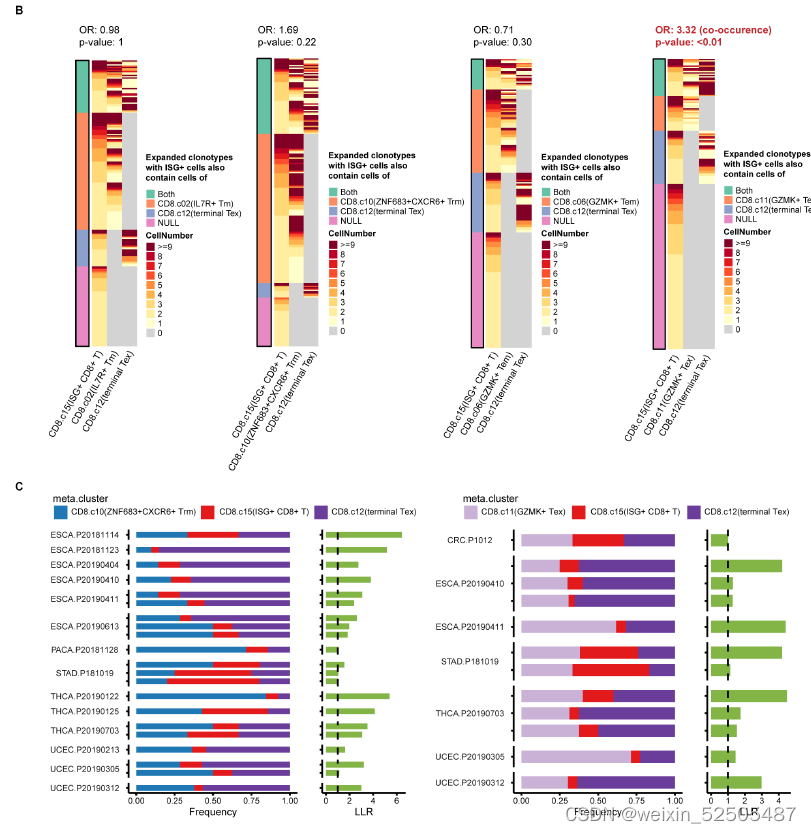
为了测试癌症类型中主要耗竭通路的prevalence,作者进一步按癌症类型对TCR sharing进行分层。在terminal Tex cells和其他metaclusters具有强TCR sharing的12种癌症类型中的11中癌症中,terminal Tex cells与GZMK+ Tex cells, ZNF683+ Trm cells, or other metaclusters in P1 or P2展示出高pTrans值。暗示了P1和P2作为作为T细胞耗竭的通用路径,作者还观察到 在某些癌症类型中,terminal Tex cells and metaclusters not in P1 or P2存在强烈的TCR sharig,包括在胰腺癌中KIR+TXK+ NK-like T cells和乳腺癌中的Tc17细胞。这些TCR sharing模式表明。除了通过Tem和Trm的共同路径以外,还存在导致T细胞衰竭的异质路径。(Fig. 2D)
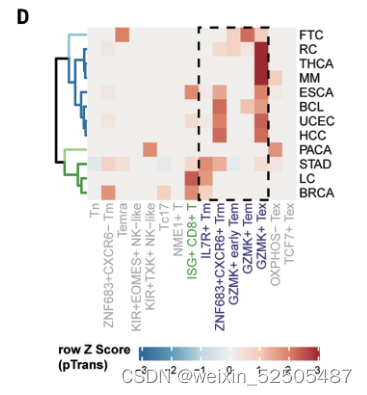
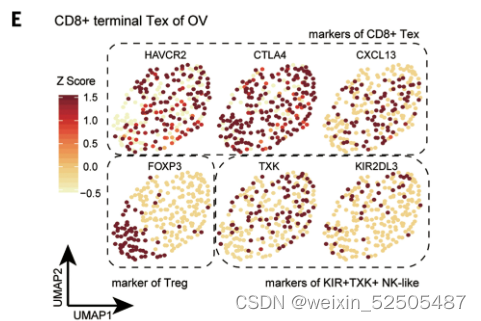
除了metaclusters之间TCR sharing以外,terminal Tex cells和其他细胞类型之间的发育联系可以通过终端Tex细胞中非耗竭状态中部分特征的signature表达来证实。卵巢癌中有一部分terminal Tex细胞表达Treg细胞主导的TF FOXP3。(Fig. 2E and fig. S20A)
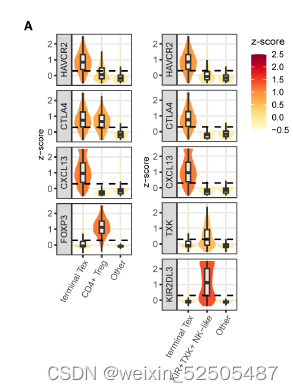
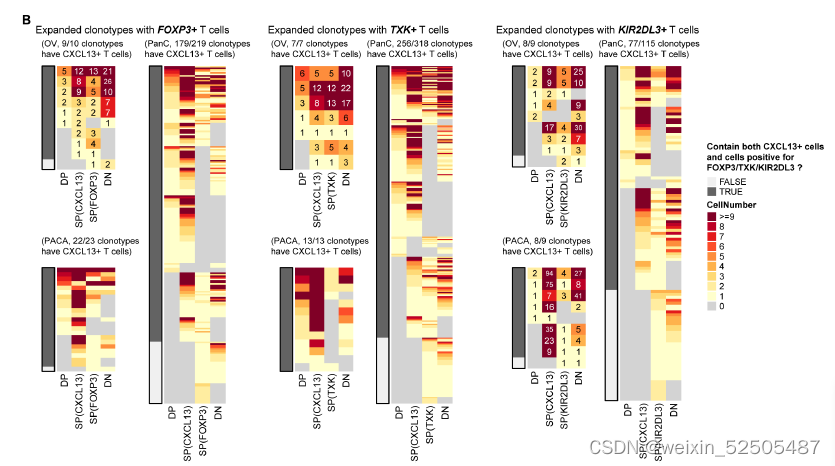
这些CD8+ FOXP3+ T细胞更趋向与与表达CXCL13的细胞共享TCR。DP, double positive, SP, single positive, DN, double negative. fig. S20B,暗示了在CD8+FOXP3+ T cells 和CXCL13-expressing cells之家intracluster transition。
同样的,在OV terminal Tex中一部分细胞类型表达KIR2DL3(killer cell immunoglobulin like receptor, two Ig domains, and long cytoplasmic tail 3)和TXK(TXK tyrosine kinase),这两个基因是KIR+TXK+ T cells的signature genes (Fig. 2E and fig. S20A),这些细胞与CXCL13表达的细胞共享TCRs fig. S20B。
除此之外,RNA velocity分析显示KIR2DL3+细胞指向CXCL13+细胞 fig. S20B。表明了KIR+TXK+ cells到耗竭细胞的转化。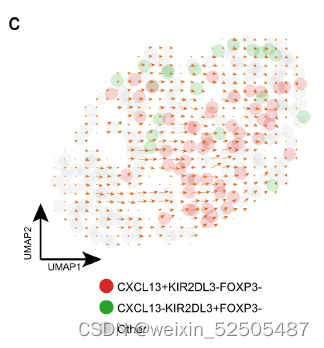
箱这样的细胞转化也可能发生在别的癌症类型中,因为在多种癌症类型中均发现表达FOXP3和KIR的细胞。fig. S20D。
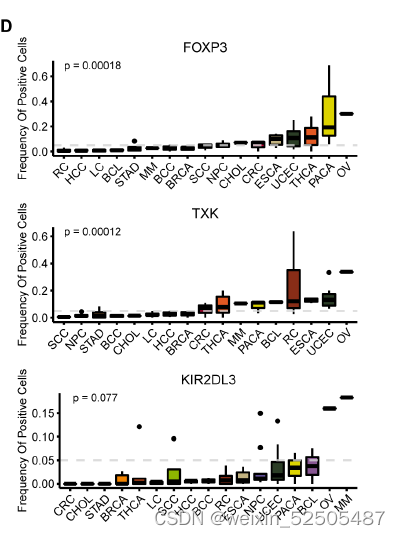
以及在多种癌症类型中发现了与Tex相似的sharing TCRs 模式,尽管其频率因癌症类型而定。fig. S20B 。
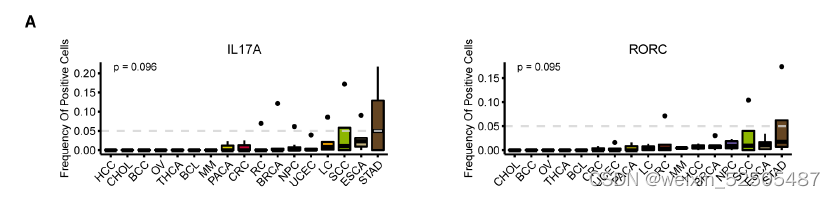
作者还进一步确认了在terminal Tex中具有癌症偏好性的基因,包括IL26,IL17A和RORC,这些基因主要表达于CD4+ TH17细胞或CD8+ Tc17细胞。作者证实了在Tc17中的表达。但是在确定的癌症类型如食管癌中,鳞状细胞癌以及胃腺癌中,这些基因也在一部分terminal Tex中表达。例如IL26。fig. S21A, Fig. 2, F and G
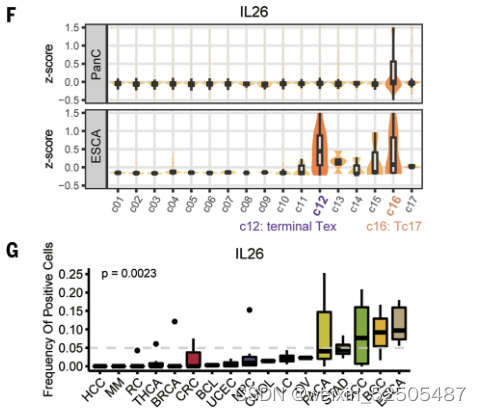
此外,包含IL26-,IL17A-,RORC-表达的Tex扩增克隆型趋向于表达CXCL13的Tex细胞 fig. S21B,暗示了表达IL26-,IL17A-,RORC-的细胞与CXCL13的细胞之间存在状态的转化。
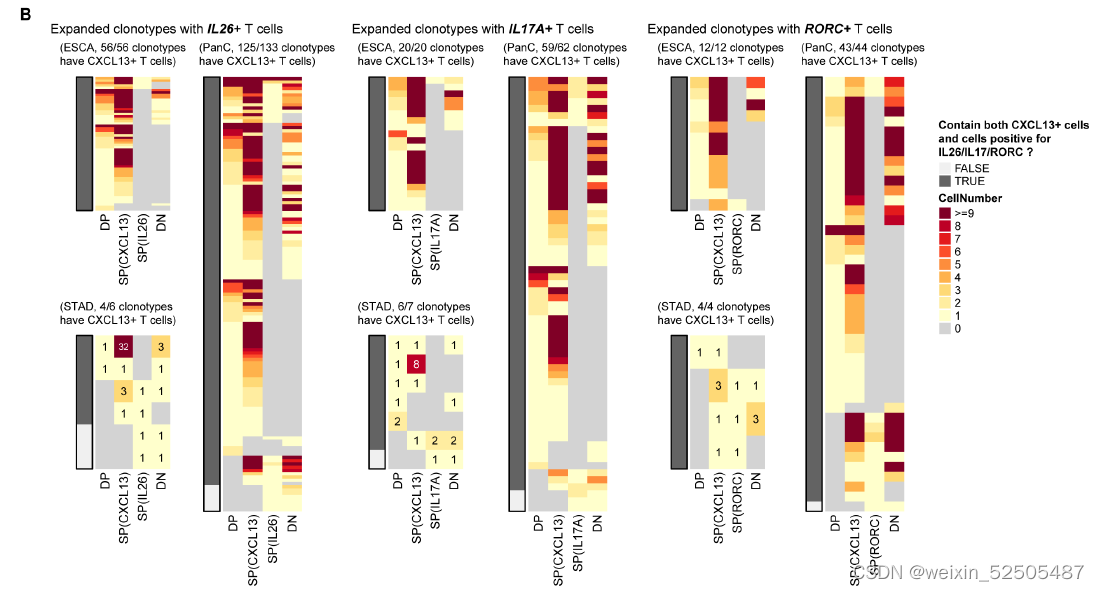
此外,一部分terminal Tex细胞表达semi-invariant α chain of MAIT,衰竭的signature以及Tc17的signature fig. S22

因此,似乎有一部分Tex具有分泌Type 17反应细胞因子的能力,这些细胞因子可能来源于Tc17细胞。有些基因优先表达,以TL26为例,它作为炎性介质,介导黏膜组织中炎性细胞因子的释放。表明了Tex在某些癌症类型中多功能的特征。
Universal and cancer type-specific transcriptional regulation of CD8+ T cell exhuaustion
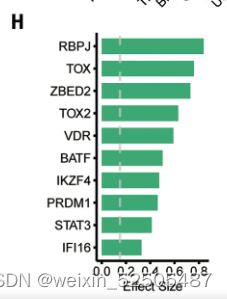
将TOX鉴定为T细胞耗竭的TF激发了人们寻找另外的TF的兴趣。作者系统的确定了与T细胞耗竭相关的TFs。满足effect size > 0.15, false discovery rate (FDR) < 0.01 by meta-analysis] in >80% of cancer types的条件的terminal Tex作为universal Tex cell regulators。TOX,TOX2,RBPJ,ZBED2,PRDM1,VDR,IKZF4,BATF,STAT3,IFI16,是按照effect size排列的前10个universal TFs。Fig. 2H, fig. S23A, and table S3
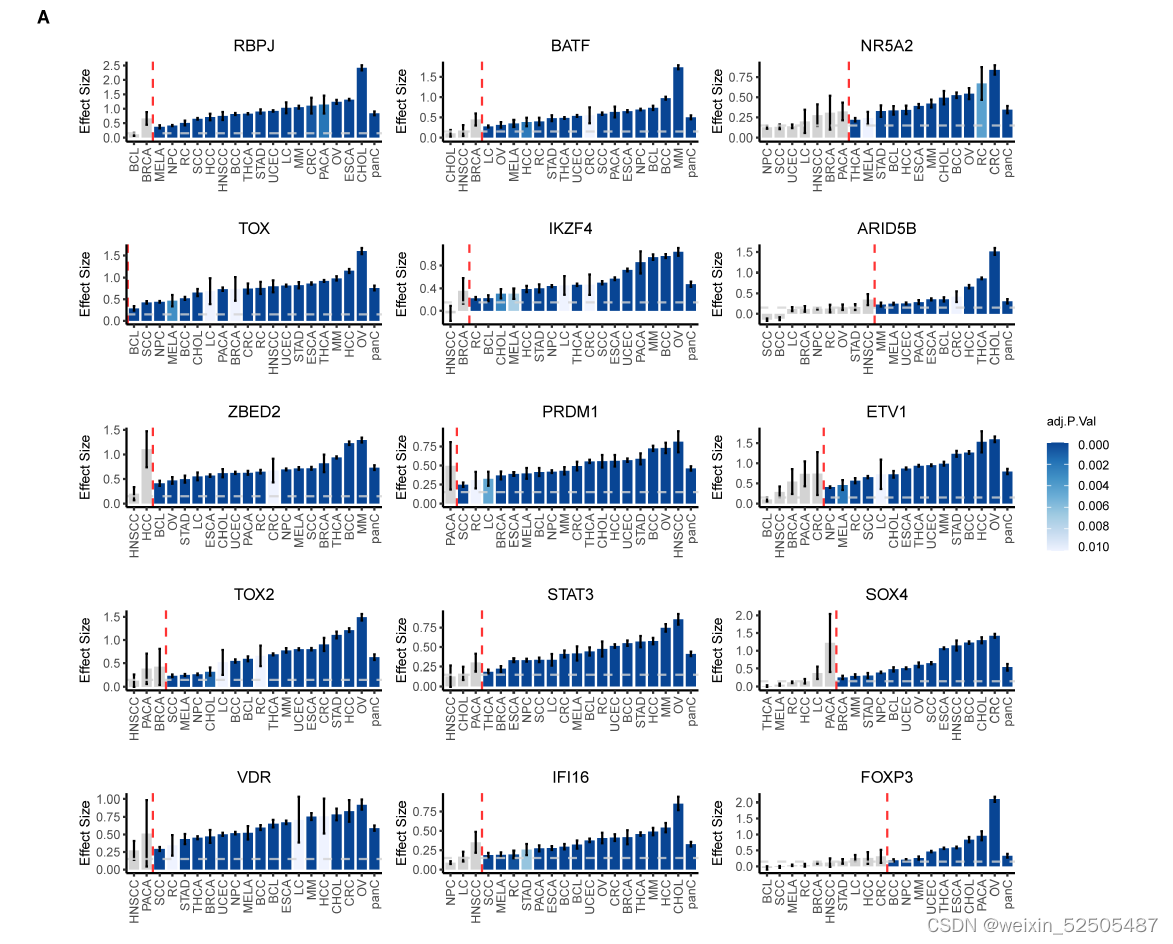
例如TOX在所有癌症类型中显示出了统计学差异。重建调节子(TF 及其靶基因)的SCENIC分析确定了靶向OX的TF,包括NR5A2,ETV1,ARID5B,其在terminal Tex中展示了高调节子的特异性并且在超过50%的癌症类型中均有统计学差异fig. S23A。Fig. 2I and table S4

支持这样的TOX的调控,使用基地细胞癌的高通量测序数据(scATAC-seq)对转座酶可及染色质的单细胞测序再分析显示,在terminal Tex中TOX的启动子或不同基因内的增强子的高可及峰值与者三个TFs相配。fig. S23B
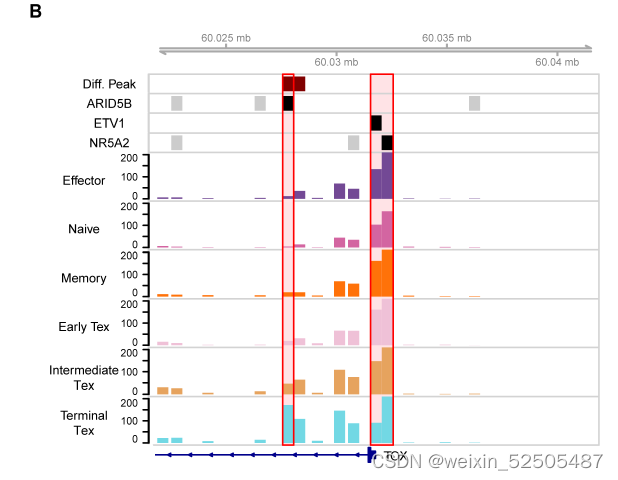
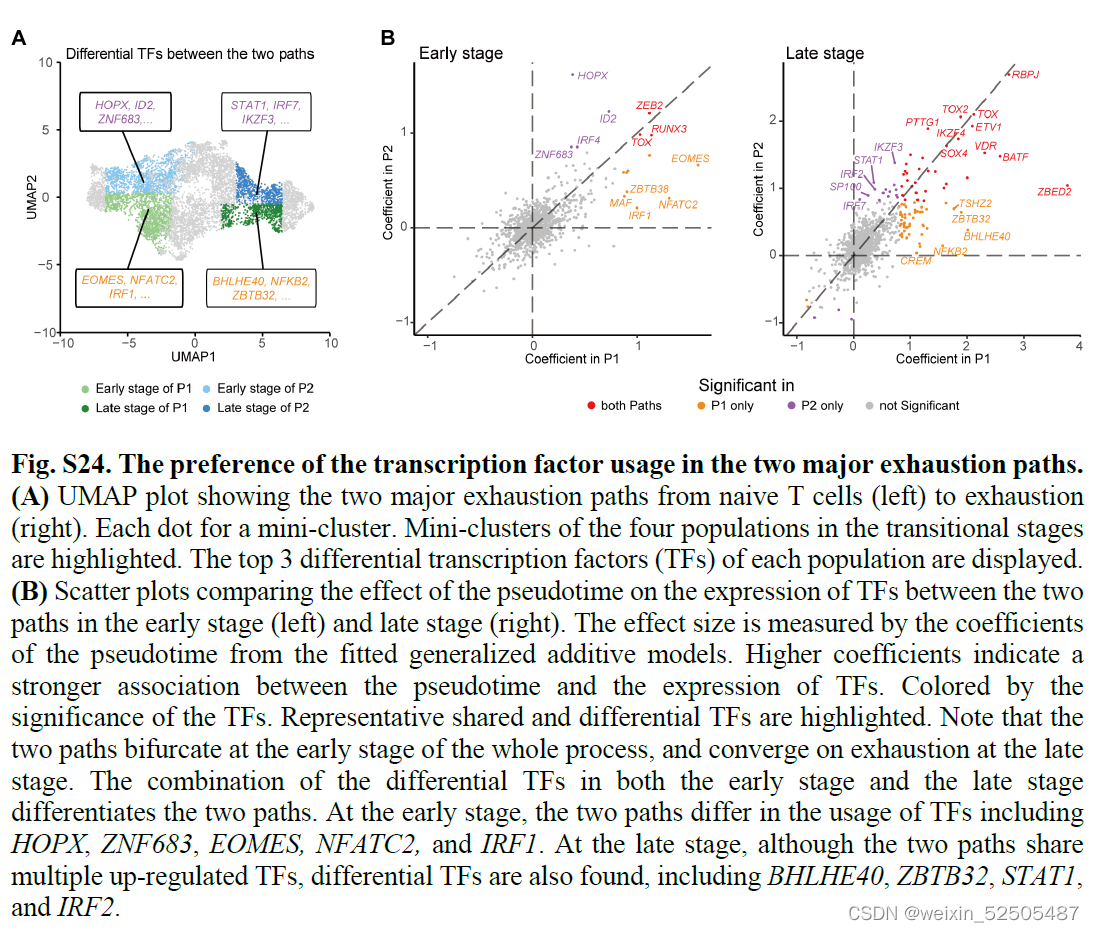
尽管Tem和Trm细胞耗竭共享多个上调的TFs,例如TOX,RBPJ和ETV1,表明这些TFs是T细胞耗竭的共同驱动力,但是不同耗竭途径在TFs的使用上可能存在不同,例如,BHLHE40和ZBTB32在P1终末阶段出现,而STAT1和IKZF3则在P2的终末阶段出现。fig. S24
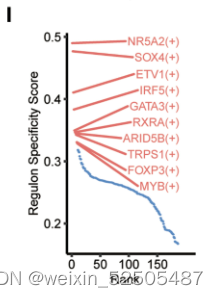
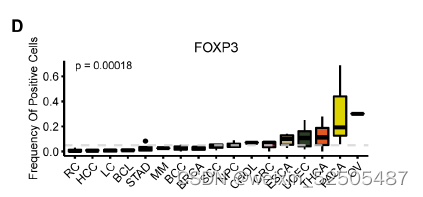
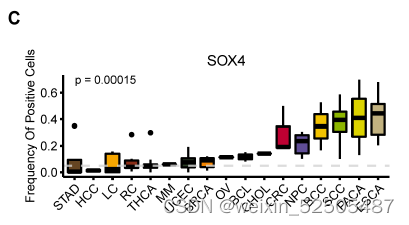
作者还确定了之前未知的和T细胞耗竭相关的转录调节因子。包括SOX4和FOXP3。特别是SOX4,具有regulon specificity score并且在所有癌症类型的2/3中具有统计学显著性。作为TGF-β下游靶标,SOX4已经被报道了在CXCL13介导的TH细胞中扮演着重要的角色并且上调Treg细胞中耗竭marker ENTPD1(CD39)的表达。因此,作者推断SOX4在Tex细胞中发挥了类似的功能,尽管这还需要进一步验证。FOXP3同样拥有高regulon specificity score,但是仅在47%的癌症中显示出统计学差异fig. S23A,因为FOXP3对于Treg细胞功能十分重要,因此,CD8+FOXP3+ T细胞(Treg细胞)和细胞耗竭之间的关系还需要进一步研究。
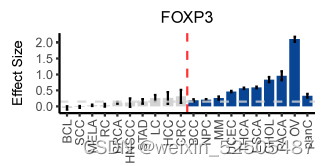
表达SOX4和FOXP3的细胞频率在癌症类型中显著不同(P < 0.01, Kruskal-Wallis tests),反映了不同TME对Tex亚型的影响。
Porperties of potientially tumor-reactive T cells in the CD4+ compartment
在CD4+ T细胞组分中,主要的肿瘤反应性的metaclusters是IFNG+ TFH/TH1和TNFRSF9+ Treg细胞。全局扩散图和RNA velocity分析揭示了CD4+ T细胞从幼稚T细胞分别发育为Temra细胞,TFH/TH1细胞或TNFRSF9+ Treg细胞**(Fig. 3A)**
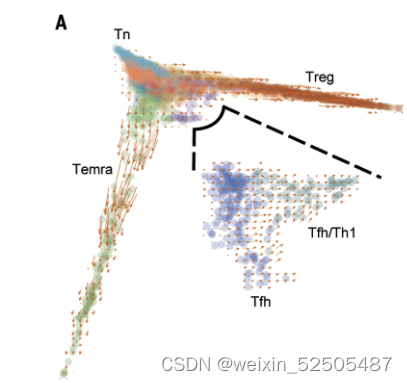
为了获得更加精细的发育轨迹,分别对每一个发育方向进行轨迹推断。两种与TFH相关的metaclusters显示出从经典的IL21+ TFH cell到IFNG+ TFH/TH1**(Fig. 3, A and B, and fig. S25, A and B)**
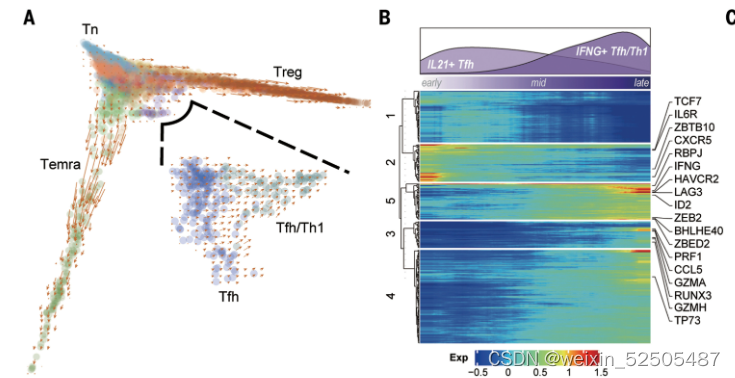
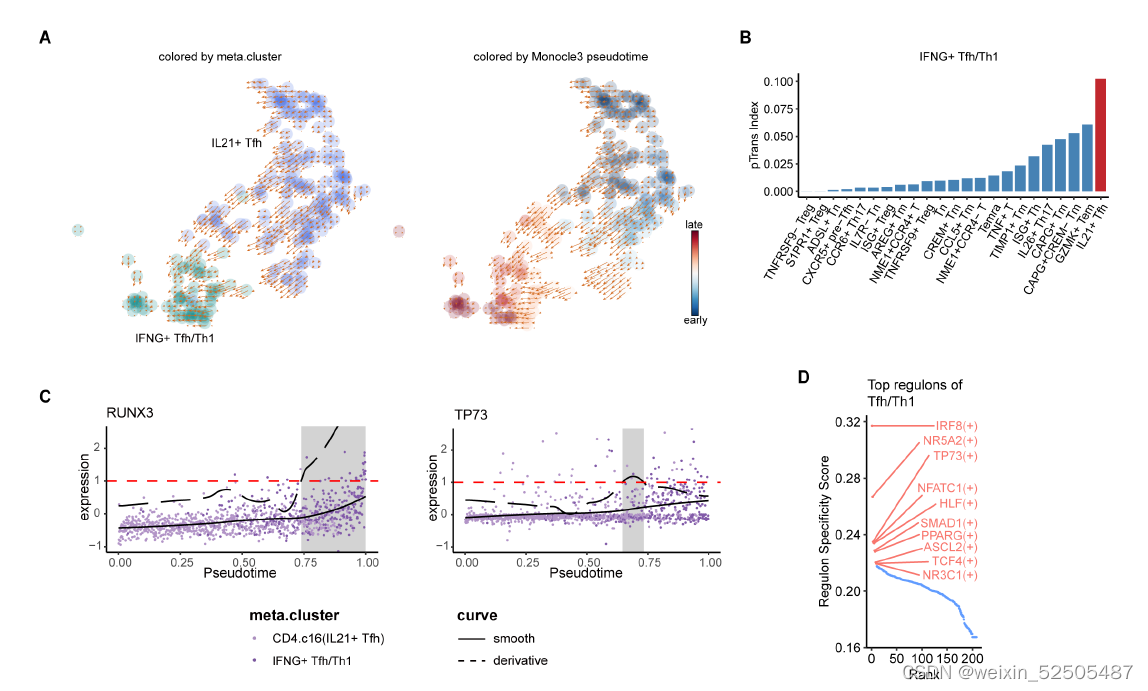
在转变的过程中,type Ⅰ反应相关的细胞因子和细胞毒效应分子包括IFNG、GZMB、PRF1显著增加,(FDR < 0.01, generalized additive model),此外,TF RUNX3在 TFH/TH1 cells中表达上调,这与之前的报道一致,即RUNX3调节CD4+ cyotoxic T细胞的细胞毒表型。TF TP73出现在较早的时间点,(fig. S25C)并被鉴定为在TFH/TH1细胞中高度特异的regulon。以上结果证明了在获取和维持TFH/TH1表型中关键的players。
对于Treg细胞,从TNFRSF9– Treg cell到TNFRSF9+ Treg cell的轨迹(Fig. 3A and fig. S26, A to D)表明从静止状态TNFRSF9–过渡到激活TNFRSF9+的状态。ISG+ Treg细胞位于轨迹的中央,表明一部人Treg细胞在激活的过程中对type Ⅰ的干扰素有反应。这与最近的报道一致,表明高ISG作为CD4+ T细胞的中间特征被激活。这种T热狗、细胞的发育轨迹在癌症类型中非常的常见在pan-cancer水平,我们没有发现从 non–Treg cell conventional TH cells所诱导的Treg细胞,但是TNFRSF9+ Treg cells在一些癌症类型中与非Treg细胞表现出一定的状态转化,例如,在B细胞淋巴瘤中TNFRSF9+ Treg cells主要与CCR6+ TH17和TFH/TH1细胞有联系,但是在子宫体部子宫内膜癌和胰腺癌则于TFH相联系,因此,各种传统的CD4+ T细胞亚群与Treg细胞有联系,但是转换的模式因癌症的类别而定。
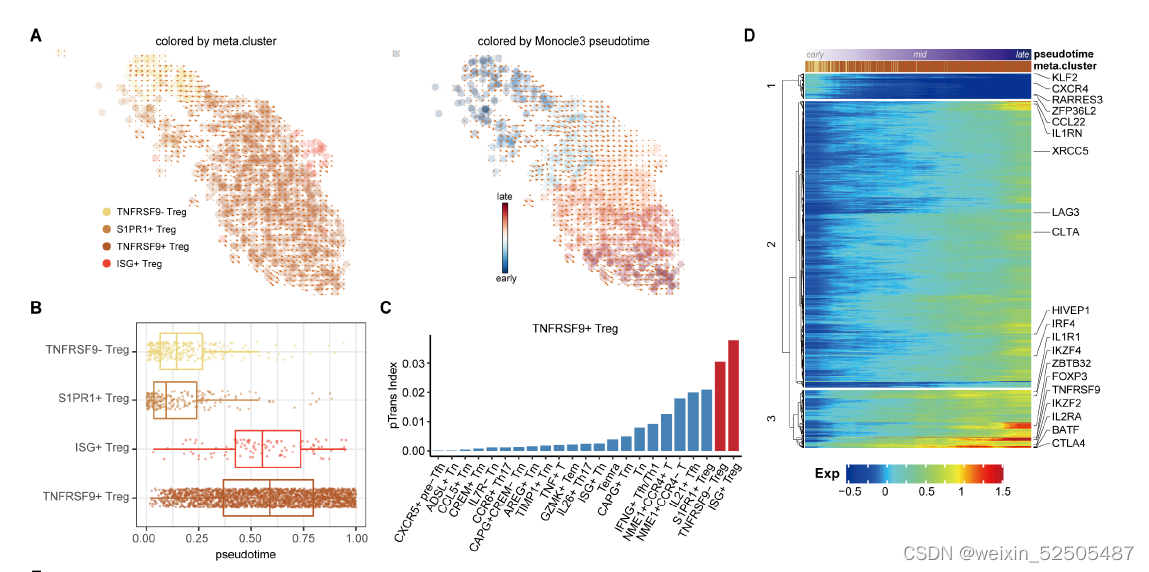
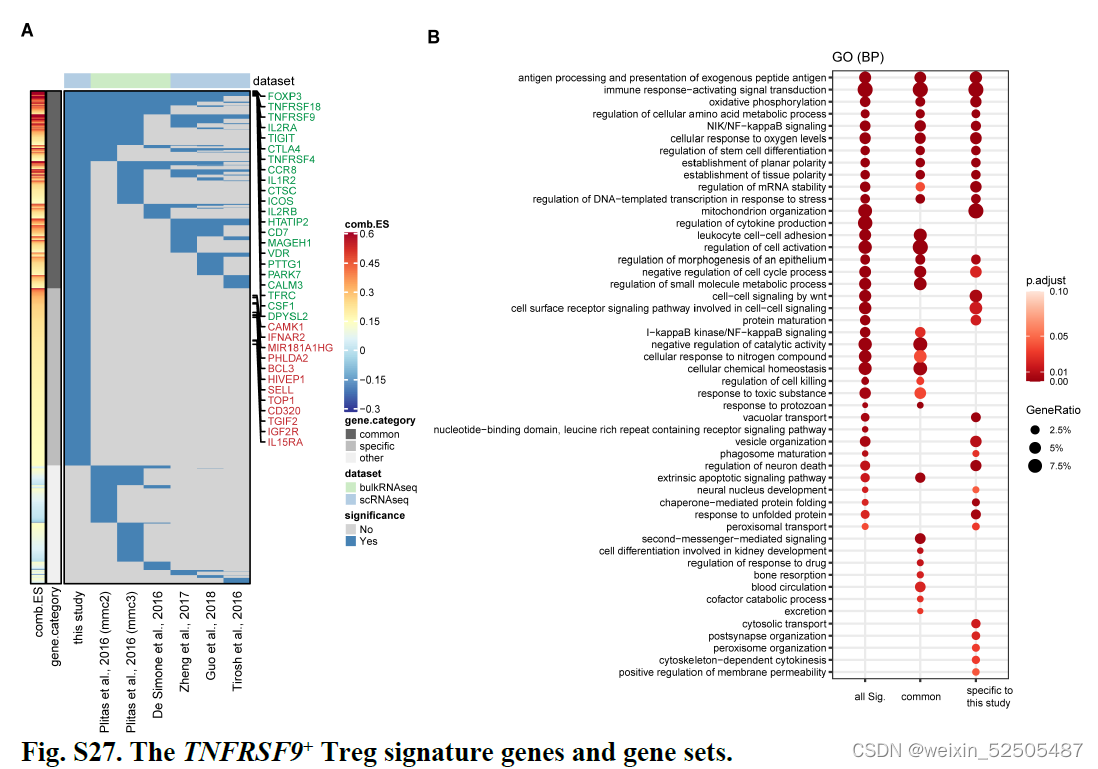
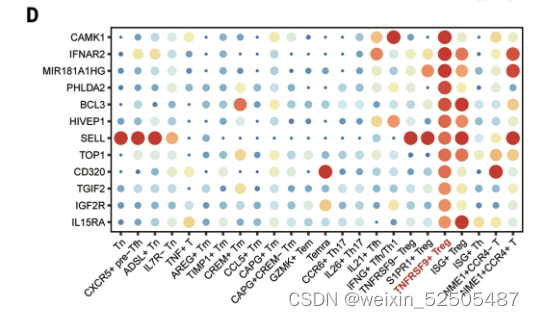
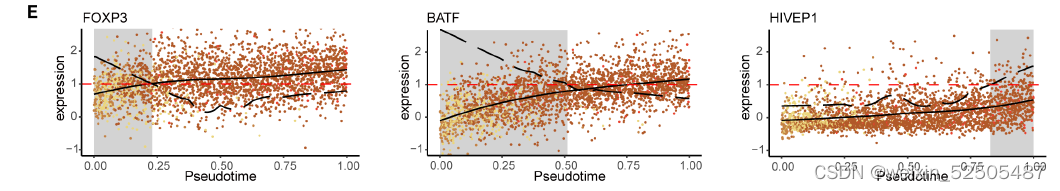
我们鉴定了之前没有报道的TNFRSF9+ Treg cell特征基因(fig. S27),包括编码具有激酶活性的膜蛋白CAMK1,IGF2R,IL15RA,IFNAR2,TOP1以及TFs:TGIF2,HIVEP1。
HIVEP1被认为是143个靶基因的关键调节因子,其中大部分也是TNFRSF9+ Treg cells的特征基因,包括TNFRSF4, TNFRSF9, ID3 (inhibitor of DNA binding 3), IL21R, and VDR (Fig. 3, E and F, and table S4)
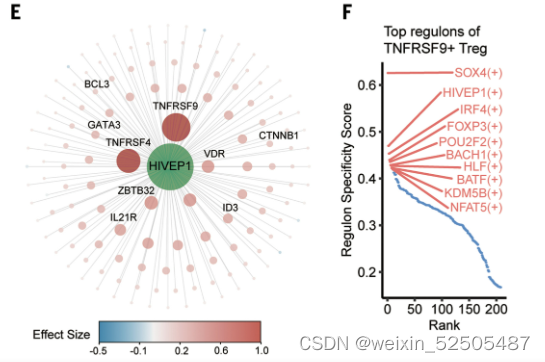
HIVEP1在Treg细胞轨迹后期加速其表达,这与其他已知的Treg细胞TFs如FOXP3和BATF在Treg细胞早期的“turned on”的模式不同。 (fig. S26E)
TME shaping the landscape of tumor-infiltrating T cells
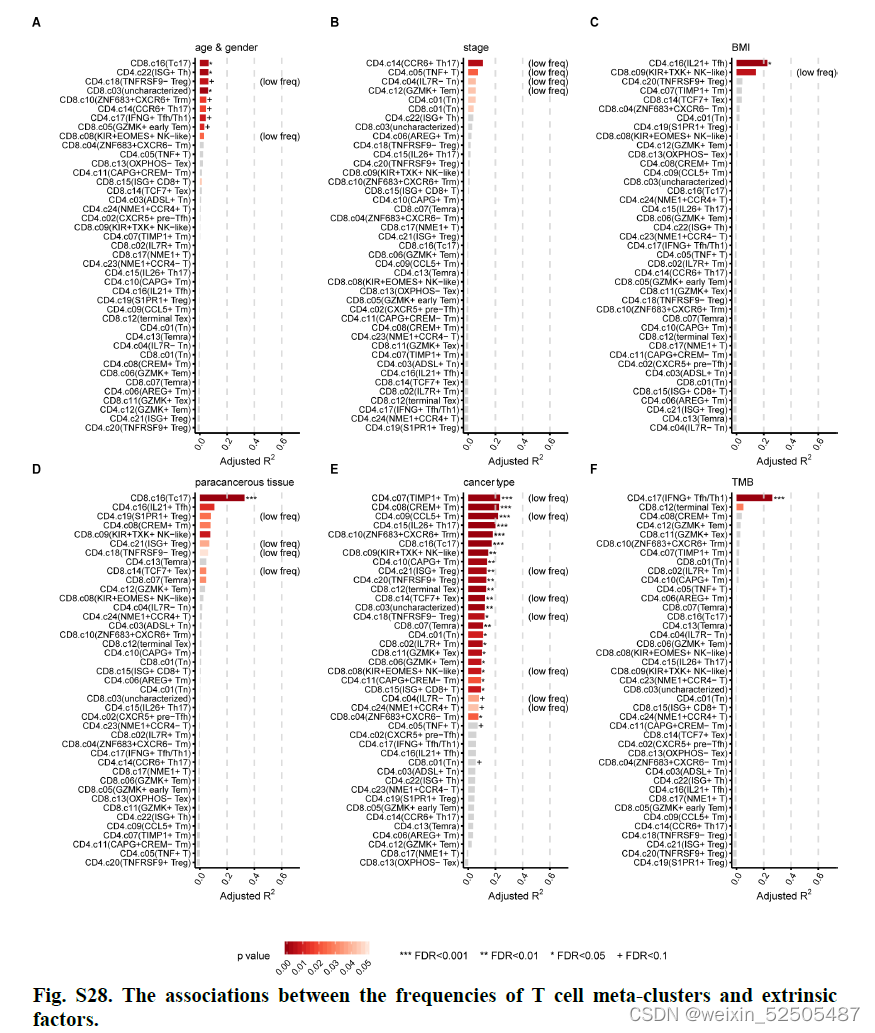
我们检查了肿瘤中不同T细胞组陈相关的外界因素,尽管没有发现每一个metacluster的细胞频率与年龄,性别以及临床stage有很强的联系**(fig. S28A S28B)。
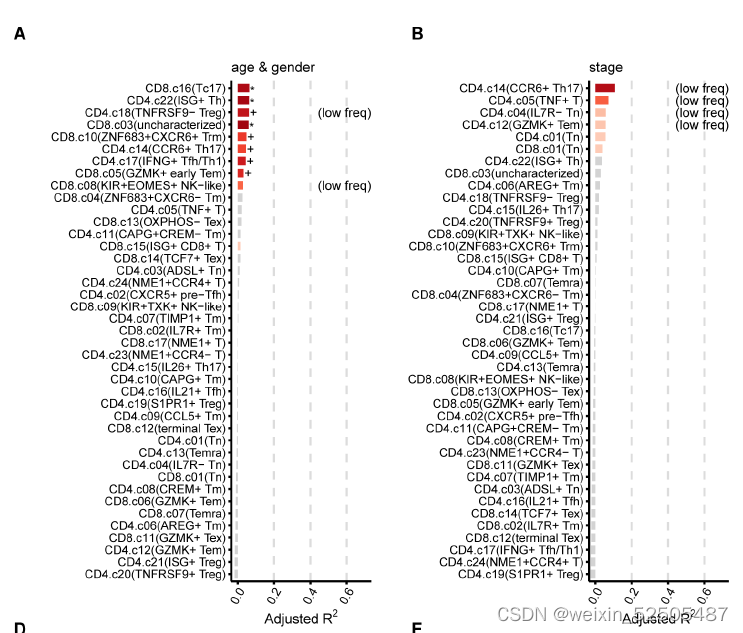
但是,我们发现了IL21+ TFH cells frequency和BMI之间存在很强的关联

除此之外,某些metacluster明显展示出组织特异性,支持宿主组织的作用,特别是肝脏富含的Tc17(figs. S28D and S29)**

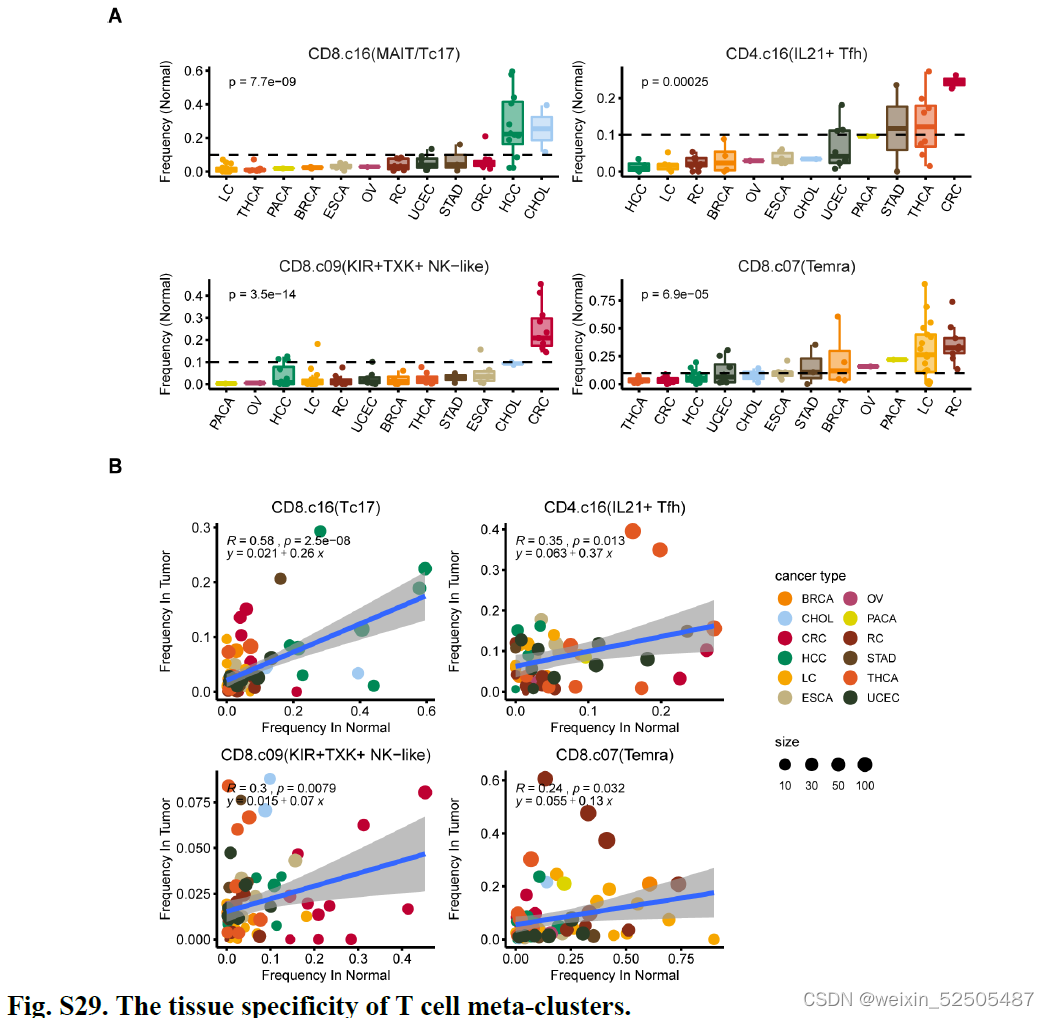
癌症类型对T细胞频率产生广泛的影响,因为观察到不同癌症类型的不同T细胞的分布模式**(Fig. 4A and figs. S28E and S30)**
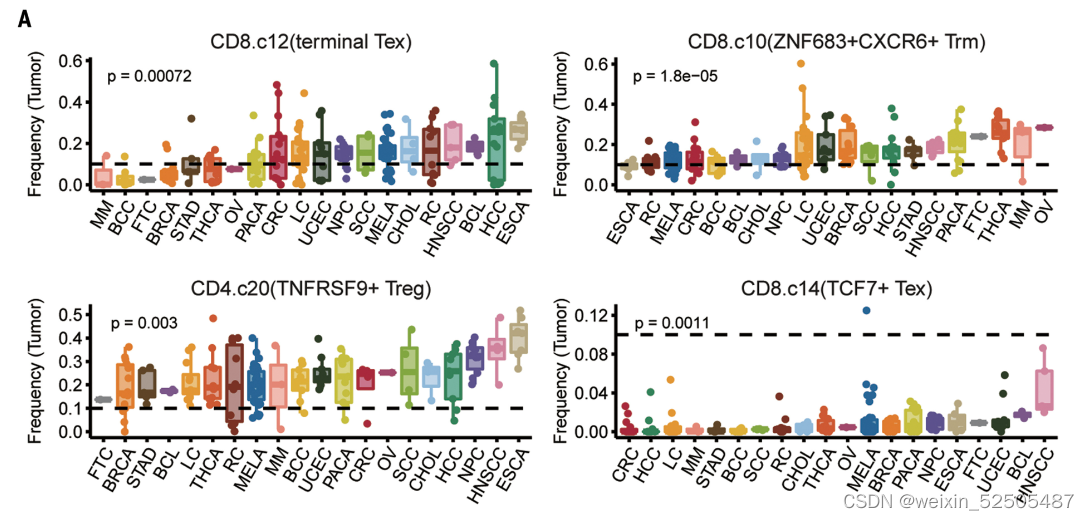
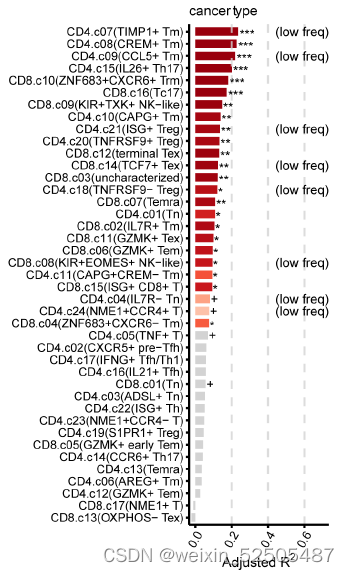
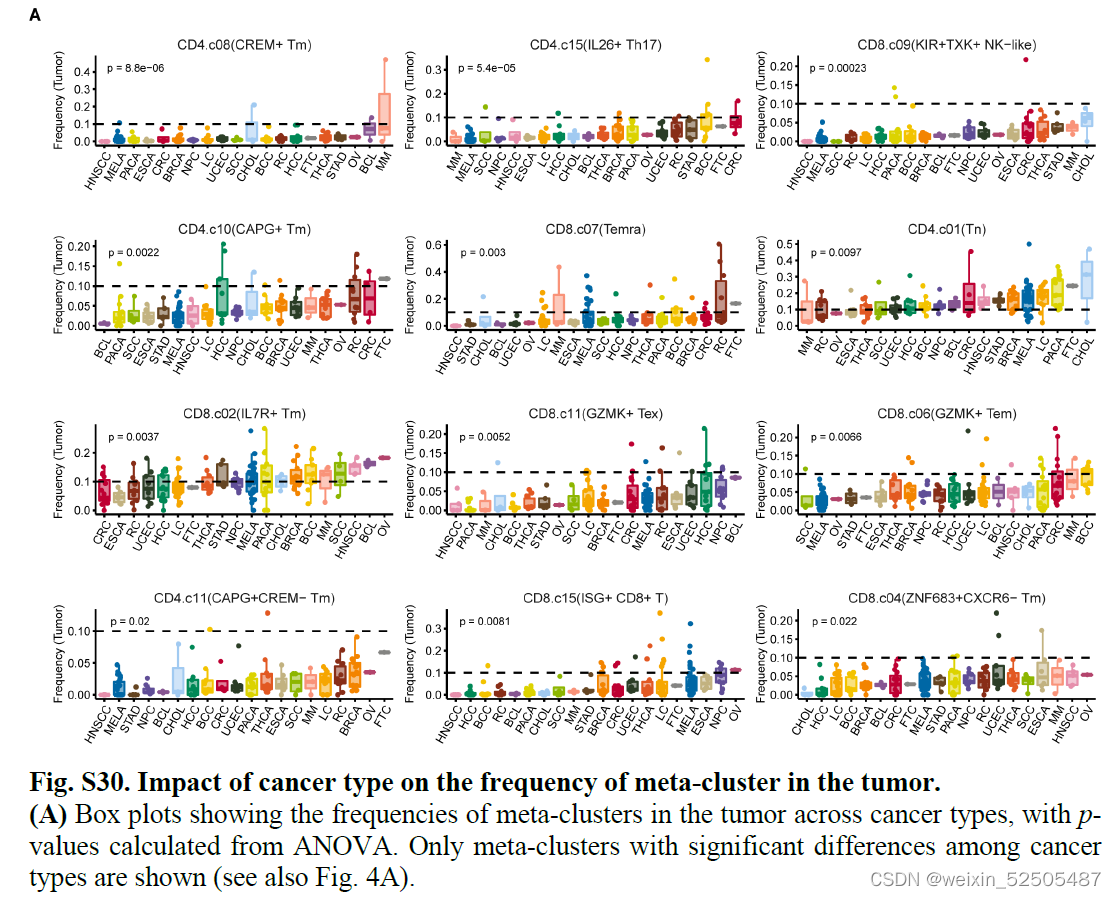
例如,terminal Tex的细胞频率从食管鳞状细胞癌的高丰度到多发性骨髓瘤的低丰度【analysis of variance (ANOVA) P = 0.00072, PVE 13.1%】,同样的,TNFRSF9+ Treg cells的中位细胞频率在所有癌症中都很高,但是在癌症类型之间的变异性也很高(ANOVA P = 0.003, PVE 13.2%),包括食管鳞状细胞癌和头颈癌显示大约高于两倍的乳腺癌和胃癌的频率。除此之外,尽管TCF7+ Tex细胞是一个罕见的亚群,但是在头颈鳞癌中显示出较高的细胞频率,但黑色素瘤样本的一个子集可以达到 12.5% 的频率。此外,尽管Temra cells在肿瘤中代表性不够,但是肺癌以及黑色素瘤的少数样本以及肾肿瘤中显示此类细胞的细胞频率要高得多。因此,针对上述细胞亚型的治疗策略应考虑不同癌症类型的变异性。
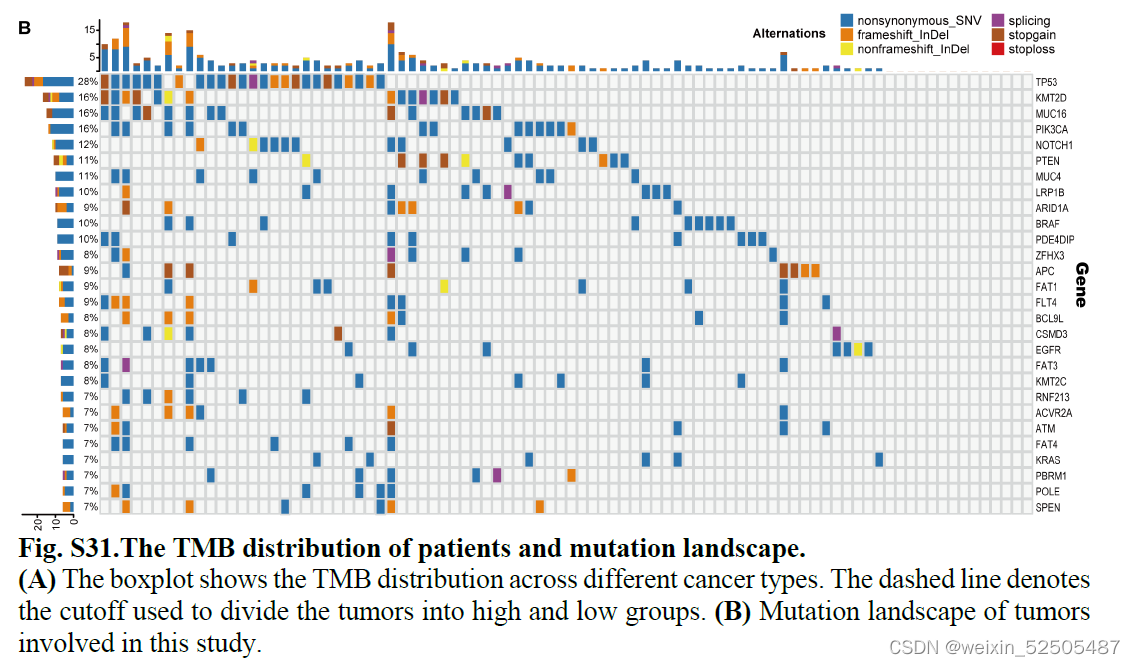
肿瘤突变负荷TMB与ICB的疗效相关。通过将肿瘤分为高低TMB组, (fig. S31A)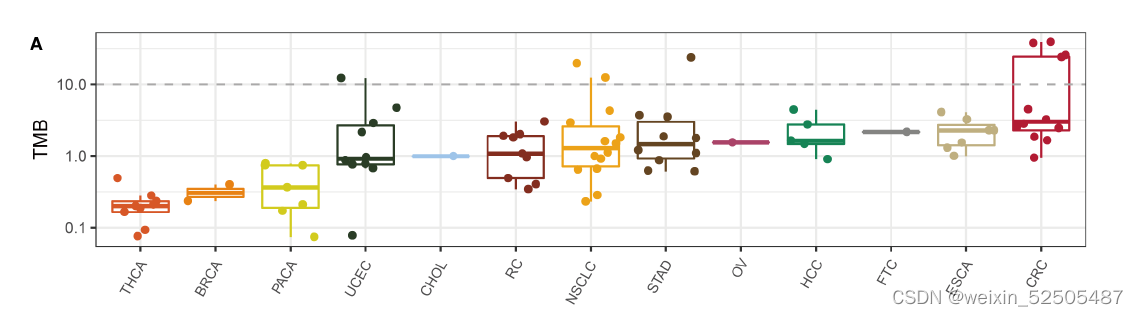
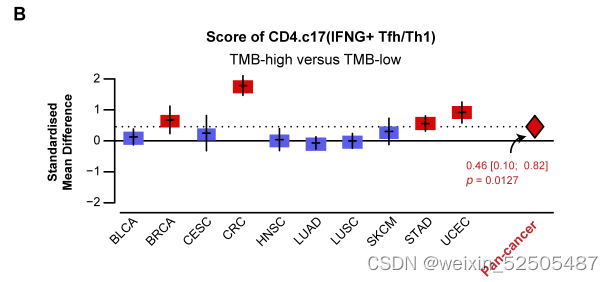
我们发现仅仅只有CD4+ TFH/TH细胞频率与TMB显示出强的相关性 (FDR < 0.001, PVE > 26.3%) (figs. S28F and S32A).

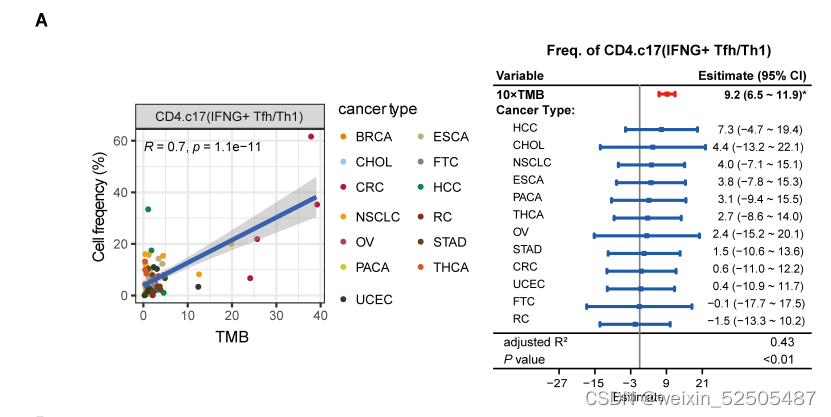
还可以使用bulk TCGA在pan-cancer和多种个体癌症类型中验证这种强联系。(fig. S32B)
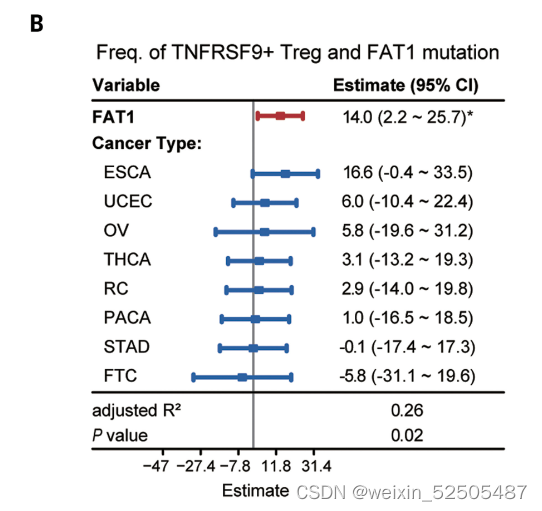
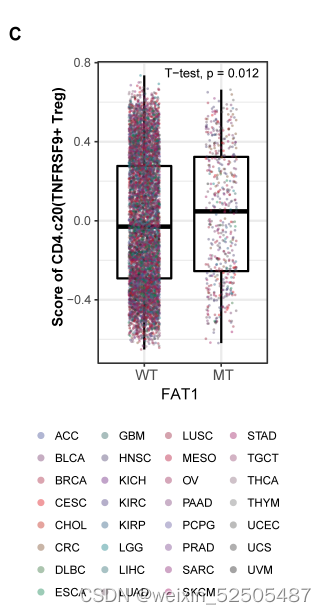
除此之外还确定了FAT1 (fatty acid translucase 1) 突变和TNFRSF9+ Treg cells之间强联系, (Fig. 4B and figs. S31B and S32C)
因此,肿瘤中T细胞的组成不仅受TMB反应的潜在新抗原数量的影响,还受癌细胞的特定体细胞突变数目的影响。
为了揭示不同癌症类型的T细胞组成的整体模式,我们检查了肿瘤中metaclusters之间的频率相关性,并确定几个高度相关的metaclusters模块**(fig. S33A)**。一个模块包含了三个metaclusters:ISG+ Treg细胞,ISG+ CD8+ T细胞,ISG+ Th细胞。而另一个模块包含四种metaclusters:terminal Tex, OXPHOS-Tex,TCF7+ Tex以及GZMK+ Tex TNFRSF9+ Treg细胞以及IFNG+ TFH/TH1细胞。具有相似基因特征但是来自于不同的compartments(CD8+ Tc, CD4+ TH, and CD4+ Treg cells)趋向于聚集在一起(for example, CD8+ Tc17 and CD4+ TH17 cells)。确定的潜在肿瘤相关性的metaclusters和某些metaclusters是负相关的,可能是由于几种机制造成的,包括metaclusters之间的动态状态转化
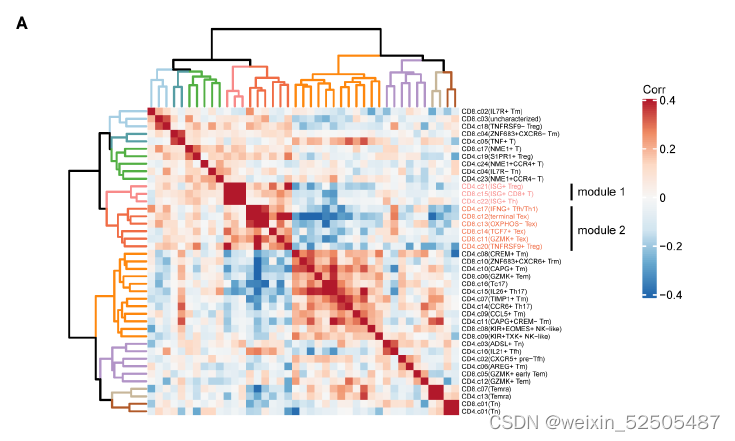
例如,CD8+ terminal Tex和CD8+ ZNF683+ Trm以及CD8+ GZMK+ Tem负相关并呈现上述状态转化的关系(aforementioned state transition relationship)。
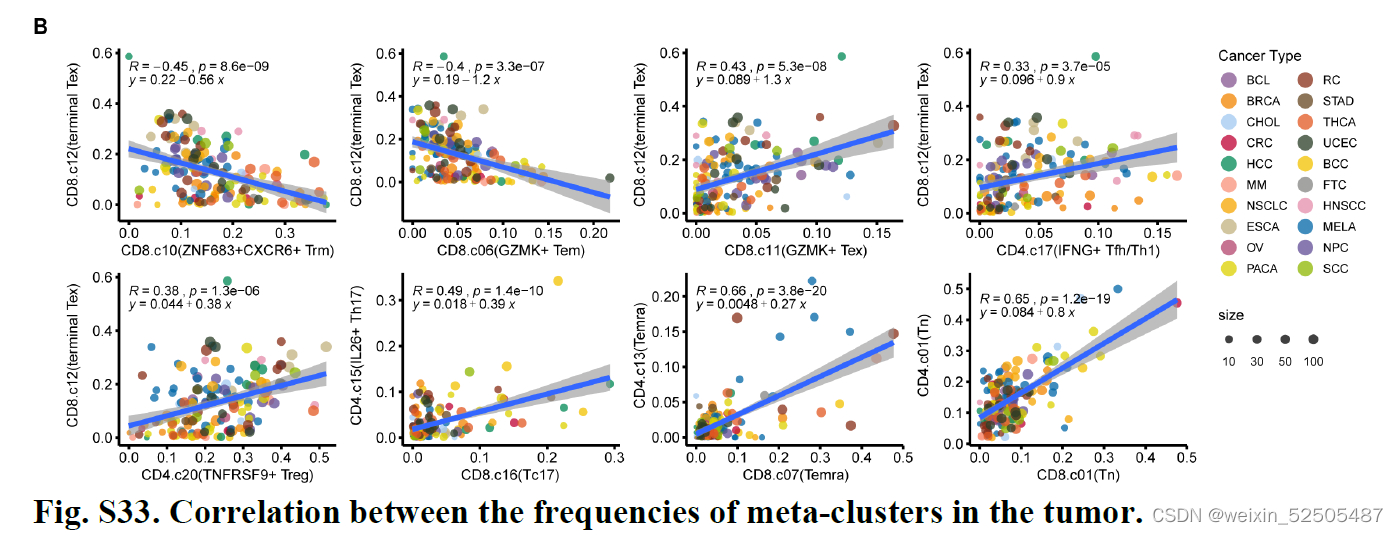
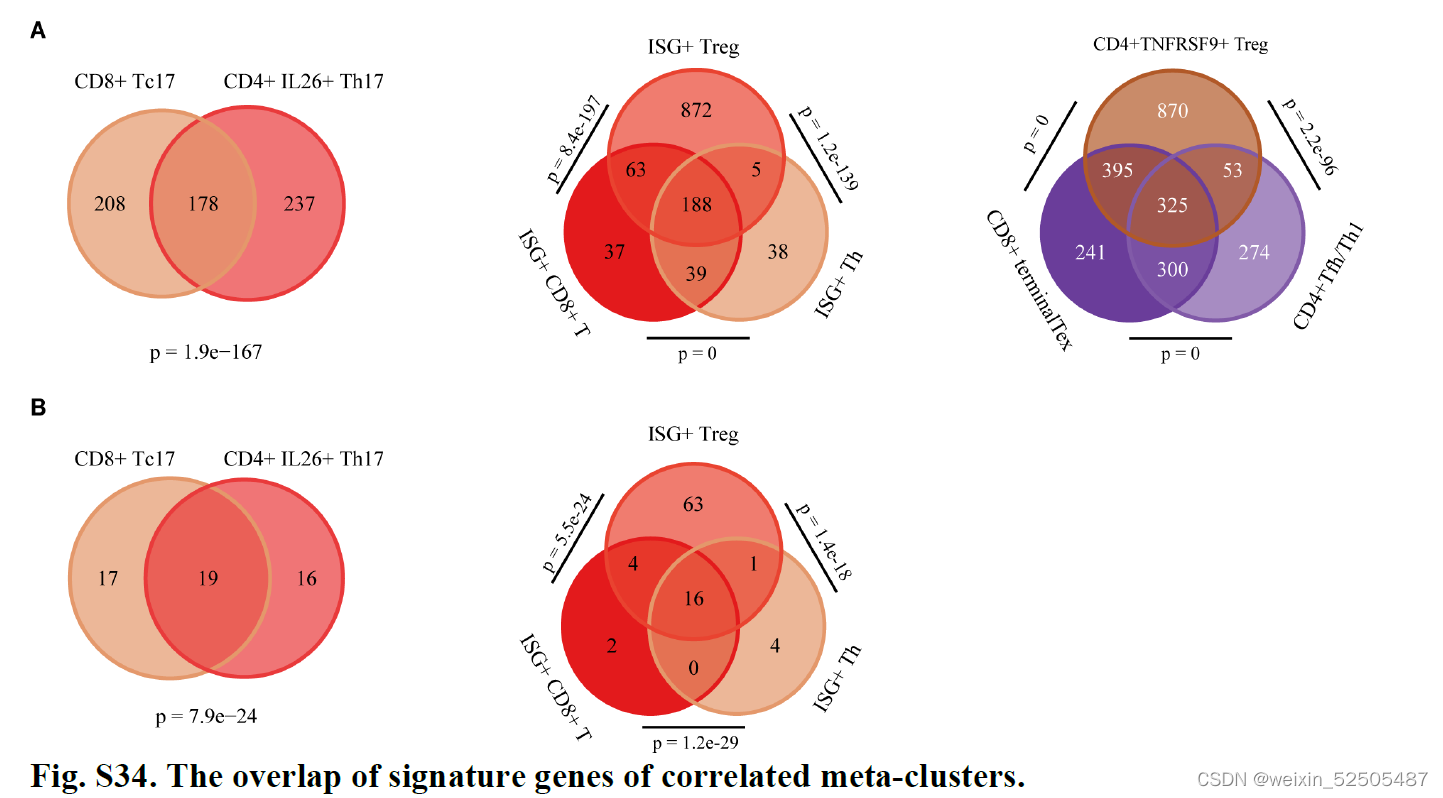
正相关的metaclusters通常在特征基因之间出现显著的重叠**(P < 0.01, hypergeometric tests) (Fig. 4C and fig. S34),表明了相同的调节子在不同的T细胞诱导了相似的转录程序。
使用重叠的signature基因和NicheNet algorithm算法去寻找共享的配体,我们发现RORC和其他type-17型反应性相关的基因在TH17和Tc17中可能被IL23A和IL15等配体诱导(fig. S35A)。

我们确定了潜在肿瘤反应性metaclusters的shared ligands,对于ISG+ metaclusters,通过NicheNet algorithm显示type Ⅰ干扰素编码的基因有可能是位于前列的ligands (fig. S35B)

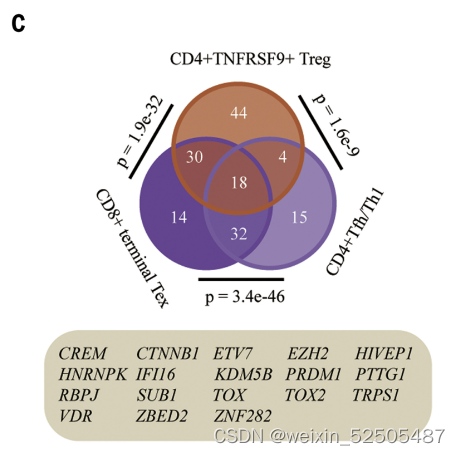
对于terminal Tex,TFH/TH1以及TNFRSF9+ Treg细胞共325个shared signature genes被鉴定,其中,18个基因 是TFs,例如,TOX, TOX2, VDR, ZBED2, ETV7, ZNF282, and HIVEP1。(Fig. 4C and fig. S34)因此,三种不同的细胞群可能利用相似的转录机制来响应TME刺激。
此外,terminal Tex和TFH/TH1 cells产生CXCL13和type Ⅰ型反应的细胞因子,TGFB1被推断为诱导两种metaclusters共享基因特征的最佳潜在配体之一(fig. S35C)**。
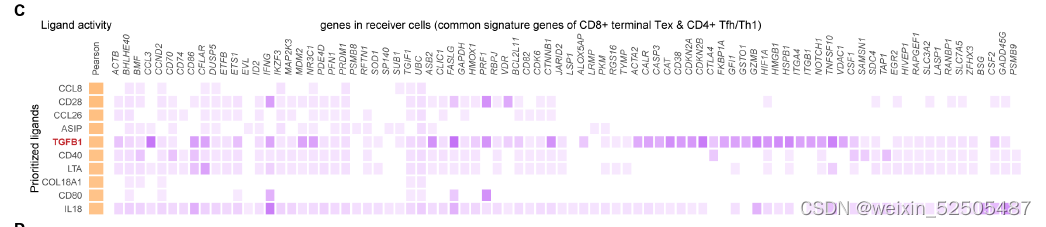
同时,TNFRSF9+ Treg细胞高表达TGF-β诱导的TFs,包括SOX4,TGIF2。此外,IFNB1被推断为介导TNFRSF9+ Treg cells and terminal Tex or TFH/TH1 cells细胞之间shared signature genes 最可能的配体之一。

上述结果支持了TGF-β and interferons可能是影响pTRTs转录程度和丰度。
Immune types of pan-cancer defined by the composition of T cells
接下来,使用这些相关的metaclusters的细胞频率,我们发现肿瘤样本可以被clustered到八组groups中(C1 to C8)(Fig. 5A)
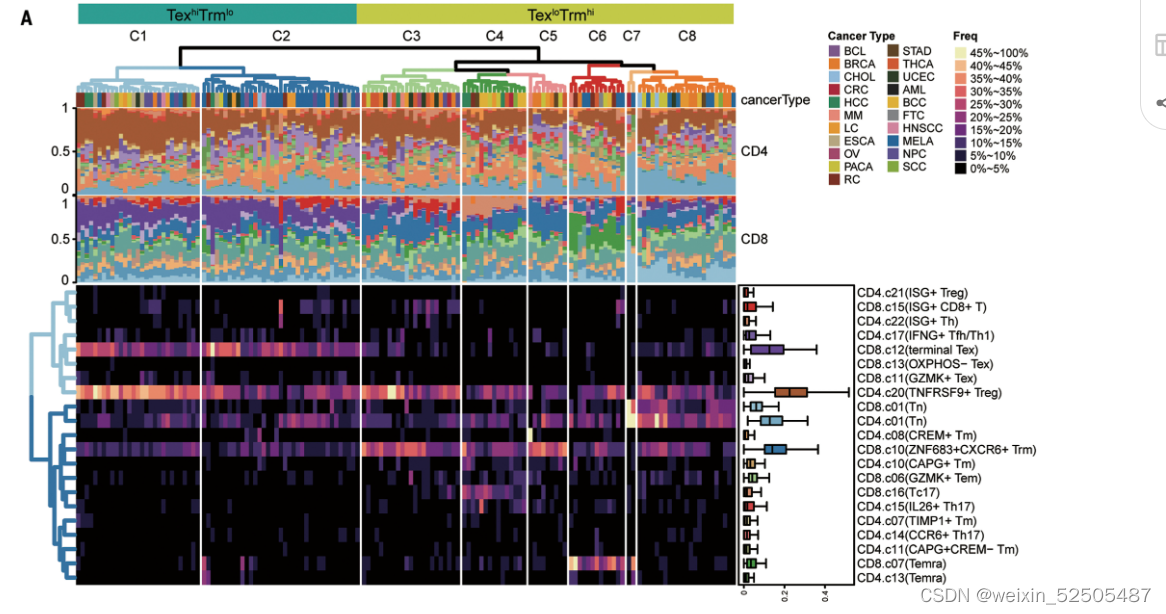
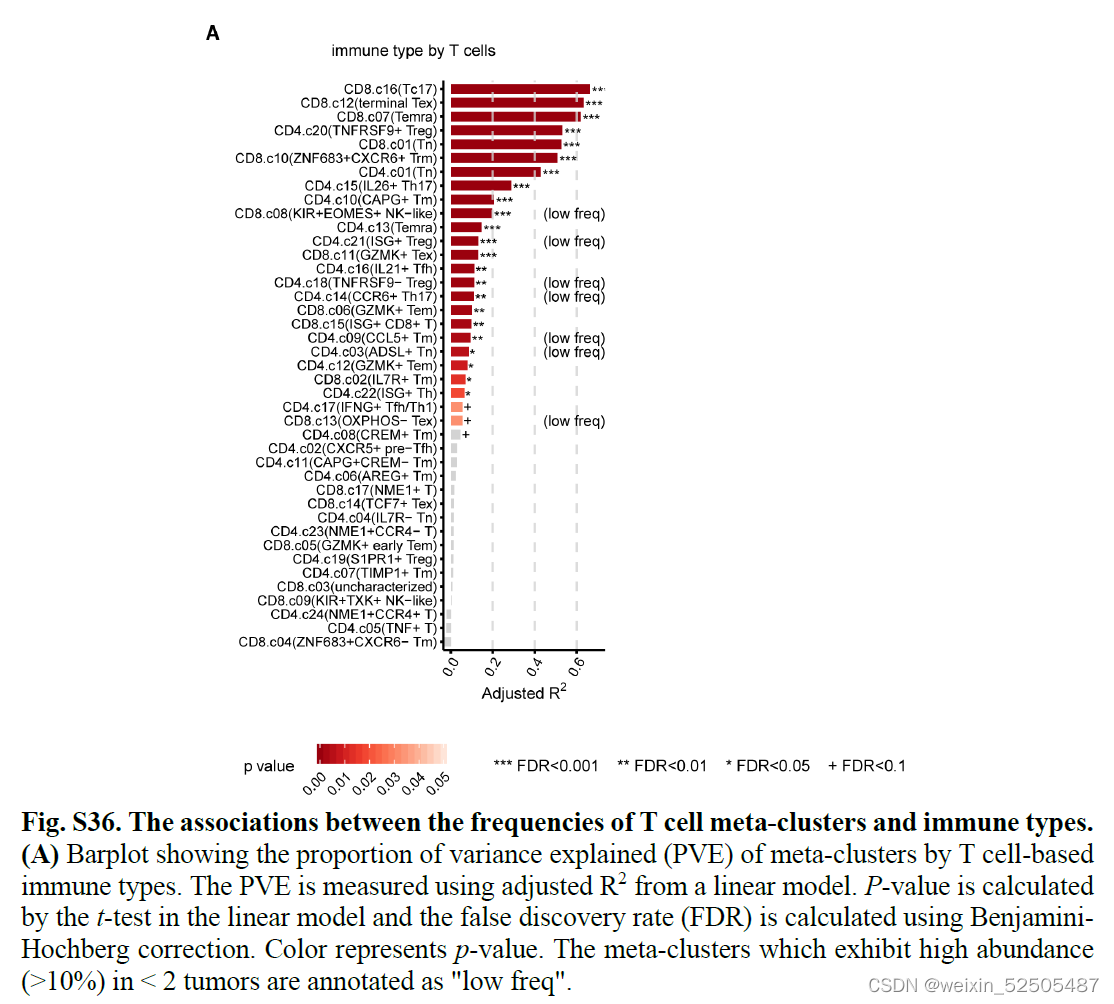
C1和C2拥有高频的terminal Tex细胞,并且C1拥有最高的TNFRSF9+ Treg细胞。C3-C8有较低的terminal Tex细胞但是具有较高的CD8+ ZNF683+ CXCR6+ Trm细胞并且可以被更进一步的分为由dominated by naive T cells (C7),enriched naive T cells(C8),enriched Temra cell(C6),enriched Tc17 or TH17(C4)以及with a low frequency of TNFRSF9+ Treg细胞(C5)。在线性模型分析的基础上,我们发现我们的分组可以解释其他因素更多的T细胞组成的变异性。(figs. S28 and S36)
尽管每一个immune type包含混合的癌症类型,但是某些确定的癌症类型具有明显的偏好。 (fig. S37)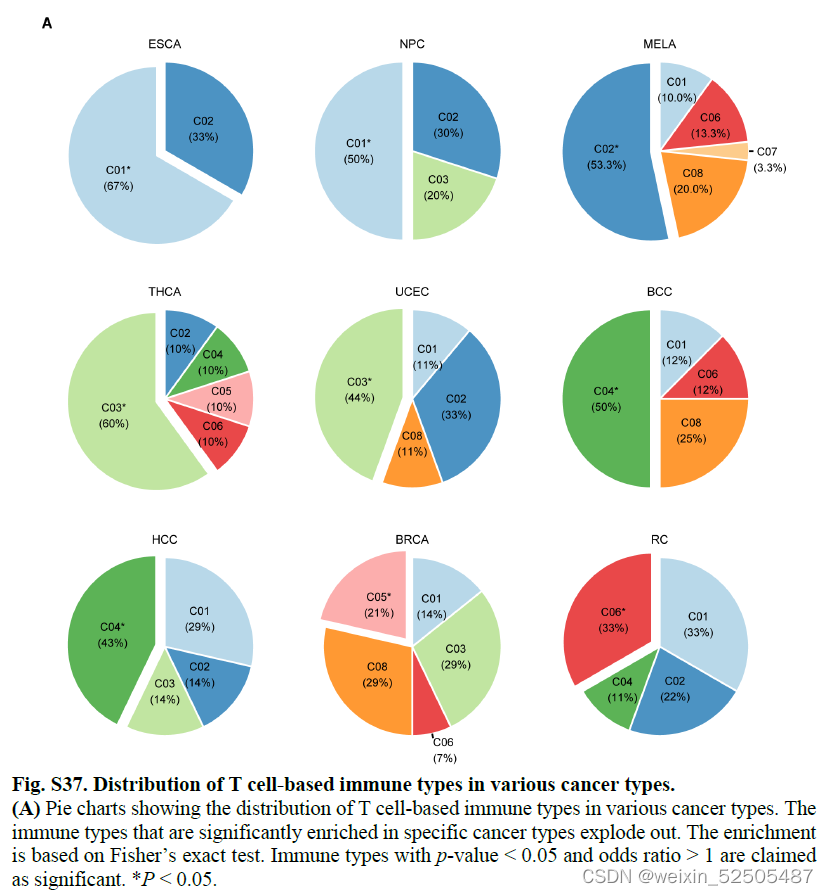
例如,接近一半的食管鳞状细胞癌都是C1 immune type。相反,甲状腺癌和子宫内膜癌富含C3 immune type。暗示了具有高T细胞抑制的癌症类型中很大一部分可能仍然受益于免疫治疗,因为ISG+ 激活T细胞的存在以及terminal Tex低丰度。超过一半黑色素瘤都是C2,主要是Tex高但是TNFRSF9+ Treg细胞低为主,这与对ICB的反应一致。基底细胞癌和肝癌都以C4 immune type为主,表明它们趋向于通过产生IL17的T细胞引起炎症反应。这些基于T细胞的免疫类型为了解肿瘤浸润性的T细胞提供了参考,这有助于指导新疗法的开发和患者的分层,而不是传统意义上癌症类型的指标
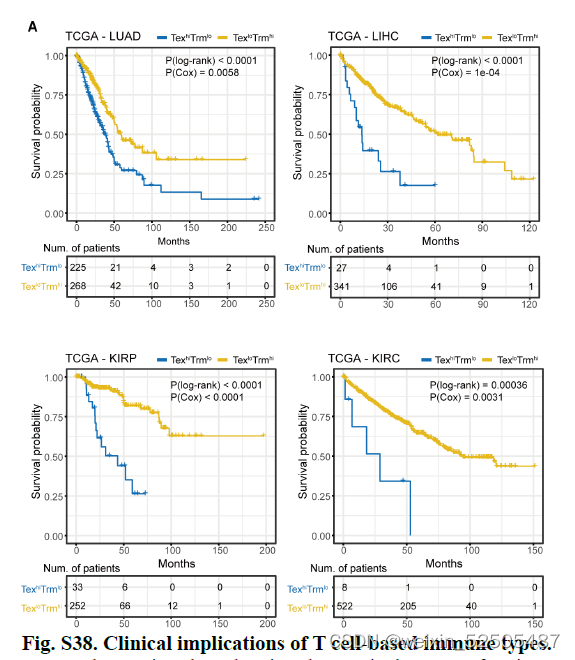
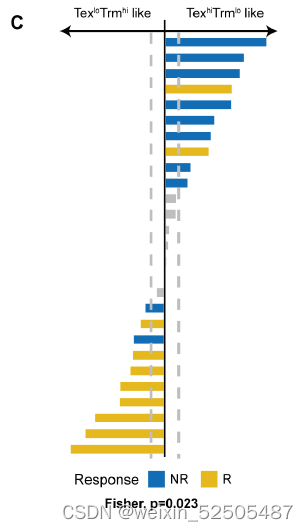
像这种免疫类型的分类可能具有临床意义。使用immune typesignatures对TCGA癌症的患者进行分层,我们发现TexloTrmhi(C3-C8)肿瘤类型趋向于比TexhiTrmlo(C1和C2)的肿瘤类型在跨癌症类型和多种individual cancer type中有更好的预后,包括肺腺癌,肝细胞癌和肾乳头状癌。Fig. 5B and fig. S38A
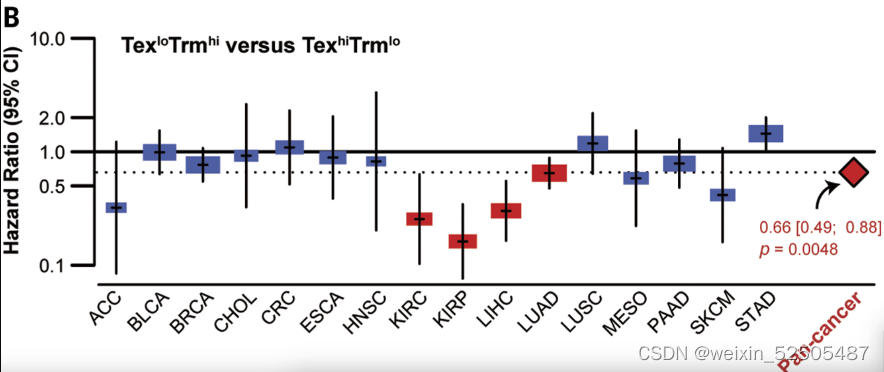
因为T细胞是大多数免疫治疗直接的靶标,基于T细胞的免疫治疗很大程度上可能与治疗的效应有关。对已经发表的PD-1抗体治疗黑色素瘤的数据重分析表明,对治疗有效应的肿瘤具有TexloTnhi(Fig. 5C and fig. S38B)
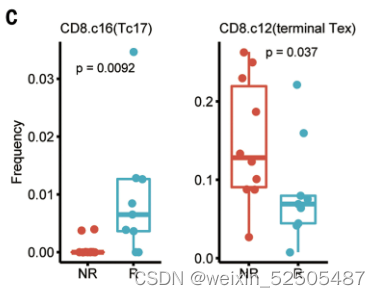
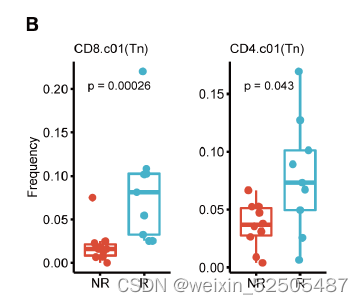
Tex细胞的效应在另一个数据集中也可以得到证明,表明这responder组的immune type为TexloTrmhi (Fisher’s exact test, P = 0.025) (fig. S38C)
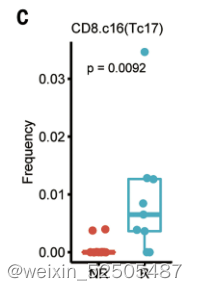
responders的治疗前肿瘤也仍然是高Tc17细胞频率,暗示了Tc17细胞在ICB治疗中的重要角色。需要进一步揭示这一发现也可能与Tc17细胞在肿瘤中耗尽相关。
Discussion
我们系统的表征了不同癌症类型中T细胞的特性,研究了从基因表达特征和异质性到细胞状态的转化和调节的不同方面。多种肿瘤富集的metaclusters-包括Tex, TFH/TH1, and TNFRSF9+ Treg cells-值得关注的是,因为它们具有潜在肿瘤的反应性
。我们的分析还揭示了T细胞耗竭的不同的途径以及这些途径的癌症类型偏好。这种landscape描绘加深了我们对癌症免疫的理解以及更好地促进治疗的发展。
在肿瘤中T细胞的状态以及浸润水平受多种因素的影响,例如肿瘤内在和代谢的因素。在我们的数据中,TMB和TFH/TH1 cells呈正相关,而BMI和TFH呈正相关。因为之前有研究显示TMB和BMI和ICB反应相关,我们的研究强调了在抗肿瘤免疫中TFH-related cell populations的重要性。除此之外,特殊的突变可能会微环境中影响T细胞的组成。FAT1突变与TNFRSF9+ Treg cells的frequency正相关。这种联系可能是由于FAT1-Hippo-YAP1的通路的功能障碍所介导。在免疫浸润细胞中肿瘤突变的影响正在显现,并且未来可能会积极探索这种联系发生的机制。此外,我们的研究还表明了不同癌症类型的TME中T细胞组成的差异并且提出了利用整体肿瘤浸润性的T细胞特性的免疫分型的方案,基于免疫typing的单细胞测序可以更好地复述这种复杂的肿瘤浸润性T细胞的特性,当发展可以个性化以获取最大临床效益时的免疫疗法时我们可以更好地informed。


